Early Vascular Plants
RHYNIOPHYTES
The earliest fossils that definitely were vascular land plants belong to Cooksonia, a genus of extinct plants (Fig. 1; Table 1). These had upright stems that were simple, short cylinders (several centimeters long) with no leaves (they had "naked stems"). They branched dichotomously, both branches being of equal size and vigor (Fig. 2). Plants of Cooksonia had an epidermis with a cuticle, a cortex of parenchyma, and a simple bundle of xylem composed of tracheids with annular secondary walls. The ends of the branches were swollen and contained the sporogenous tissue. Unlike the sporangia of algae, these were large and multicellular. Only the central cells were sporogenous and were surrounded by several layers of sterile cells; the sporangium wall had to open to release the spores. The plants were homosporous; there were no separate microspores and megaspores. Fossils that have these general characters are called rhyniophytes.
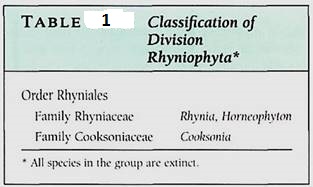
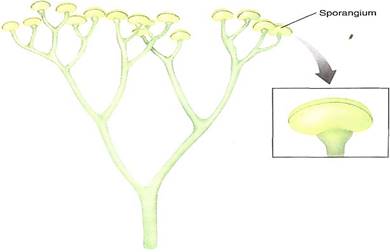
FIGURE 1:A reconstruction of Cooksonia caledonica, the earliest known plant that had xylem—tracheids with annular secondary walls. Important features are its sporangia at the ends of branches, the lack of leaves, and the dichotomous branching.
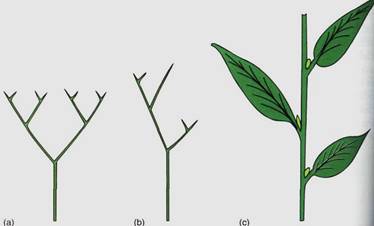
FIGURE 2:Branching patterns. (a) If a stem forks, resulting in two equal stems, it is a dichotomy or dichotomous branching. (b) It is pseudomonopodial branching if one stem is definitely lager and tends to form a trunk. (c) If one stem dominates the system absolutely, it is monopodial branching, as in most seed plants.
Rhynia was another rhyniophyte, an early vascular plant similar to Cooksonia (Fig. 3). It had a prostrate rhizome, upright naked stems, and terminal sporangia. It had stomata and guard cells in the epidermis and a layer that appears to have been a cuticle.

FIGURE 3: Reconstruction of Rhynia major (a) and Rhynia gwynne-vaughanii (b). They strongly resemble Cooksonia, and we know that they definitely had rhizomes, upright stems, and rhizoids. Inset shows the sporangium cut away, revealing spores.
Xylem Structure of Early Vascular Plants. Early vascular plants had two types of organization of xylem. In both the center is a solid mass of xylem with no pith; this is a protostele. In an endarch protostele, protoxylem is located in the center and metaxylem differentiates on the outer edge of the xylem mass. The other type of stele present in early vascular plants is an exarch protostele, with metaxylem located in the center of the xylem mass and protoxylem on the edges as several groups next to the phloem. Another type of stele, which did not evolve until later, is the siphonostele, one in which pith is present in the center, as occurs in the stems of flowering plants. Because xylem fem is often preserved well in fossils, its characteristics of exarch/endarch and protostele/ siphonostele are usually available for study.
The xylem in many specimens of Rhynia is well preserved: It was a round cylinder without pith but with protoxylem in the center and metaxylem on the exterior—that is, an endarch protostele (Fig. 4). All xylem cells were tracheids with annular thickenings (Fig. 5). Around the xylem was a layer of phloem-like cells, then a parenchymatous cortex and epidermis.

FIGURE 4:Early vascular plants had two types of organization of xylem. In both, the center is a solid mass of xylem with no pith. This is a protostele, common in roots of almost all plants. (a) In an endarch protostele, protoxylem is located in the center, and metaxylem is on the periphery. Endarch protosteles occurred in the rhyniophytes. (b) In an exarch protostele, metaxylem is located interior to the xylem mass and protoxylem on the edges, as several groups next to the phloem. Exarch protosteles are found in fossils of zosterophyllophytes.

FIGURE 5: A tracheid with annular secondary walls found in Rhynia gwynne-vaughanii (X 200). (Courtesy of K. ]. Niklas, Cornell University)
Life Cycles of Early Vascular Plants. In the same rocks with Rhynia are other fossils of similar plants. Two of these, Lyonophyton and Sciadophyton, were gametophytes, not sporophytes. The ends of the stems bore flattened cup-shaped areas that contained gametangia, both antheridia and archegonia, but not sporangia. These plants had upright, dichotomously branched stems, vascular tissue with tracheids, stomata, and a cuticle.
The nature of these early gametophytes is important in understanding the early evolution of vascular plants. The discovery of an alternation of heteromorphic generations in algae was possible because the individuals could be grown in culture and their life cycle could be observed. This is impossible with fossils, but it is possible to suspect that because Rhynia-type sporophytes occur together with Sciadophyton-type gametophytes, they might have been alternate phases of the same species. If this is true, these plants had an alternation of isomorphic generations, and later evolution into the seed plants involved reduction of the gametophyte to just a few cells and elaboration of the sporophyte into a more complex plant. This hypothesis is the transformation hypothesis (Fig. 6). An alternative hypothesis, the interpolation hypothesis, postulates that the earliest plants were monobiontic without a multicellular sporophyte (Fig. 7). A small sporophyte was presumed to come into existence when a zygote germinated mitotically instead of meiotically. The sporophyte generation would have gradually evolved in complexity while the gametophyte generation remained small. A sporophyte generation would be inserted (interpolated) into the monobiontic life cycle. In this hypothesis, nonvascular plants such as mosses are thought to be intermediates in the progression from green algae to vascular plants (Fig. 7). We have no way to decide which hypothesis more accurately approximates the actual origin of terrestrial sporophytes. The presence of Horneophyton, Lyonophyton, and Sciadophyton supports the transformation hypothesis, as does the similarity between Anthoceros and Horneophyton. But in mosses and liverworts, several species with the most complex capsules, which are parts of the sporophyte phase, seem to be the most derived species. Perhaps mosses and liverworts arose separately from green algae by interpolation. At present, data are insufficient to be certain.
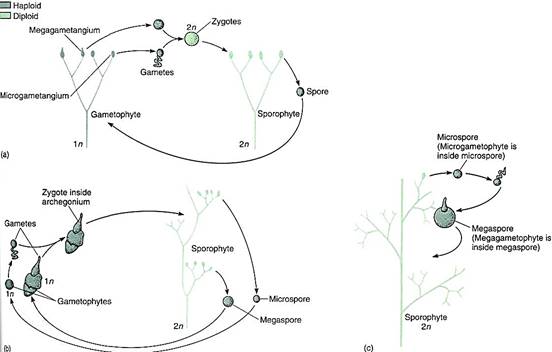
FIGURE 23.7:The transformation theory of the origin of the vascular plant life cycle postulates that in early land plants (a), gametophytes were upright and dichotomously branched, with epidermis, cuticle, and vascular tissue, just like sporophytes. (b) With time, sporophytes became larger and more complex and gametophytes became simpler. Even though these are two plants of the same species, different phenotypes are adaptive for each, because each contributes to the survival of the plant in different ways In the species illustrated here, the gametophytes have become so small that the microgametophyte develops within the spore wall and the megagametophyte protrudes from the spore only slightly. (c) With continued reduction, it is possible—but neither necessary nor inevitable—for the megaspore and its megagametophyte to be retained inside the megasporangium and remain on the parental sporophyte, an important step in the process of seed evolution.
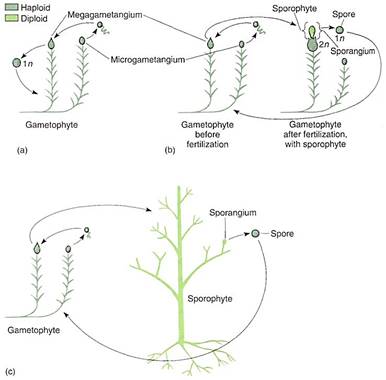
FIGURE 7:The interpolation theory. (a) The very earliest land plants were postulated to have no sporophyte; instead the zygote "germinated" by meiosis. (b) At a later stage in evolution, the zygote would germinate mitotically and produce a simple sporophyte that in the early stages would have consisted of a sporangium and perhaps also a foot. (c) With continued evolution, the sporophyte would have become progressively more elaborate while the gametophytes became simpler.
ZOSTEROPHYLLOPHYTES
Another group of early vascular plants are the zosterophyllophytes, also called the zosterophylls, named after the principal genus Zosterophyllum (Fig. 8a; Table 2). They were small herbs without secondary growth. Many of their features were similar to those of rhyniophytes, but three characteristics make us think they are a distinct group: Their sporangia were lateral, not terminal; the sporangia opened transversely along the top edge (Fig. 8b); and their xylem was an exarch protostele, that is, protoxylem on the outer margin and metaxylem in the center (Table 3).
Although these distinctions may seem minor, we are certain that in all the more recent plants with large leaves (ferns, conifers, flowering plants), sporangia are terminal rather than lateral; thus, plants like Rhynia may have been the transitions between algae and later seed plants, but Zosterophyllum was not. Instead, some of the simplest vascular plants alive today, the lycophytes, have lateral sporangia, and they may represent a line of evolution based on Zosterophyllum-like ancestors (Fig. 9).



FIGURE 8: (a) Zosterophyllum rehenanum plants were quite similar to those of the rhyniophytes, but the ends of fertile branches bore numerous lateral sporangia, not a single terminal one. Also, sporangia (b) opened by a suture that passed over the top of the sporangium, not up its side.
FIGURE 9:the very earliest vascular plants—those that immediately preceded Cooksonia—are unknown. Out of that group came rhyniophytes and zosterophyllophytes. We are not sure whether they evolved as two completely separate lines or as one line that soon diverged into one group with terminal sporangia and endarch protoxylem and a second line with lateral sporangia and exarch protoxylem. Nor do we know whether the nonvascular plants evolved out of these very earliest vascular plants or directly from green algae.
Zosterophyllum is a genus of extinct plants that grew as small bunches, only about 15 cm high. They probably grew in swampy, marshy areas: Upper portions of their stems had cuticle, ordinary epidermal cells, and stomata, but lower portions did not, presumably because they grew under water. Stems of Zosterophyllum were naked (smooth), branched dichotomously, and contained a small amount of xylem that consisted of tracheids with annular and scalariform secondary walls. Sporangial walls were several layers thick, and all spores were the same size, so they must have been homosporous.
The zosterophyllophytes and rhyniophytes shared so many characters that we believe they must have evolved from a common ancestor. They represent two lines of evolution, the rhyniophytes giving rise later to the ancestors of seed plants and ferns, and the zosterophyllophytes giving rise to the lycophytes. (See the section on the microphyll line of evolution.)Other genera of zosterophyllophytes show that significant morphological changes evolved quickly. In Rebuchia the sporangia occurred together on the ends of specialized branches (Fig. 10). In Crenaticaulis some branching was pseudomonopodial: Larger, trunk-like shoots bore smaller, shorter lateral shoots. These are the morphological bases for producing cones and trunks. Several of the zosterophyllophytes had a smooth surface (see Fig. 8), but others had outgrowths of tissue called enations that ranged from quite small to long, thin scales. Enations increased the photosynthetic surface area of the plants, and in Asteroxylon they contained stomata and a small trace of vascular tissue that ran from the stele through the cortex to the base of the enation (Fig. 11).

FIGURE 10:This reconstruction of the zosterophyllophyte Rebuchia ovata shows several significant advances in body construction. The stems that bore sporangia were specialized, keeping the sporangia close together and elevated into the wind; other stems branched repeatedly, forming a large photosynthetic surface. Some branches were larger than others, so branching was no longer strictly equal dichotomies. These derived features are not in the line that led to terns and seed plants; analogous evolutionary changes had to occur in that line as well.

FIGURE 11:Reconstruction of Asteroxylon, an early lycophyte, showing the surface covered with enations, small flaps of photosynthetic tissue. In species without enations, stems were round and oriented vertically, not very good for harvesting sunlight.





















