آخر المواضيع المضافة

 الفاكهة والاشجار المثمرة
نخيل التمر
الحمضيات
الاشجار ذات النواة الحجرية
المحاصيل
المحاصيل البقولية
محاصيل الالياف
محاصيل زيتية
محاصيل الحبوب
محاصيل الخضر
القرعيات
المحاصيل المنبهة و المحاصيل المخدرة
المحاصيل السكرية
نباتات الزينة والنباتات الطبية والعطرية
الحشرات النافعة
النحل
دودة القز(الحرير)
تقنيات زراعية
التصنيع الزراعي
الانتاج الحيواني
الطيور الداجنة
الاسماك
الابقار والجاموس
الاغنام
آفات وامراض النبات وطرق مكافحتها
الحشرات
امراض النبات ومسبباتها
الحشائش والنباتات الضارة
الفاكهة والاشجار المثمرة
نخيل التمر
الحمضيات
الاشجار ذات النواة الحجرية
المحاصيل
المحاصيل البقولية
محاصيل الالياف
محاصيل زيتية
محاصيل الحبوب
محاصيل الخضر
القرعيات
المحاصيل المنبهة و المحاصيل المخدرة
المحاصيل السكرية
نباتات الزينة والنباتات الطبية والعطرية
الحشرات النافعة
النحل
دودة القز(الحرير)
تقنيات زراعية
التصنيع الزراعي
الانتاج الحيواني
الطيور الداجنة
الاسماك
الابقار والجاموس
الاغنام
آفات وامراض النبات وطرق مكافحتها
الحشرات
امراض النبات ومسبباتها
الحشائش والنباتات الضارة | ماهية ونشوء الفايروسات |


|
|
أقرأ أيضاً
التاريخ: 21-6-2018
التاريخ: 26-6-2018
التاريخ: 2-7-2018
التاريخ: 16-8-2017
|
ماهية ونشوء الفايروسات
أثارت الفايروسات النباتية والأمراض التي تسببها اهتمام الباحثين بعد الدراسات الرائدة التي أجريت في نهاية القرن التاسع عشر ونتيجة لصغر حجم الفايروسات وعدم إمكانية رؤيتها بالمجاهر الضوئية المتوفرة وقتئذ وكذلك لاستحالة عزلها وتنميتها على بيئات زراعية خارج النسيج النباتي الحي مما يسهل دراستها كالفطريات والبكتريا، فقد فشل الباحثون في تلك الفترة المبكرة من نشوء علم الفايروسات في معرفة ماهية هذه الكائنات لذلك وضعت العديد من الفرضيات لتفسير ماهية هذا الكائن والتي تأثرت بالمكتشفات الحديثة من الطفيليات وخاصة البكتريا باعتبارها أصغر الكائنات الدقيقة المكتشفة حينها حيث بحث واضعوا هذه الفرضيات عن تقديم تفسير علمي لأسباب هذه الأمراض بما كان متوفرا عندهم من إمكانات علمية وعملية إلا أن كل هذه الفرضيات سقطت اليوم ولم تعد مقبولة علميا وذلك بعد التقدم العلمي الكبير الذي مر به علم الفايروسات والذي كشف الكثير من أسرار هذا الكائن الغريب.
الفرضيات السابقة التي وضعت لتفسير ماهية الفايروسات النباتية
وضعت عدة فرضيات لتفسير ماهية فايروسات النبات في بواكير نشوء علم الفايروسات وجميعها لم تعد مقبولة اليوم وهي:
(1) الفرضية الإنزيمية وهي التي اقترحها "وود" Wood سنة 1899 مفترضا أن الفايروس هو إنزيم غريب يفرزه طفيل ما وبنى فرضيته استنادا على ملاحظته بزيادة كمية الإنزيمات المؤكسدة في خلايا نباتات التبغ المصابة بفايروس موزائيك التبغ TMV)).
(2)الفرضية البكتيرية وهي التي تبناها ماير و ايفانوفسكي في نهاية القرن التاسع عشر بعد انجازهما لبحوثهما على فايروس موزائيك التبغ مفترضين أن المرض الذي يسببه هذا الفايروس يعود إلى بكتيريا صغيرة جدا تعبر المرشحات البكتيرية ولا تشاهد بالمجهر الضوئي.
(3) فرضية البروتوزوا (الحيوان الأولي) والتي اقترحها "ايفانوفسكي" سنة 1903 معتقدا أن الأمراض الفايروسية تسببها أنواع من البروتوزوا نتيجة رؤيته للأجسام الضامة المسماة "الأجسام السينية" X -bodies التي شاهدها بالمجهر الضوئي في الخلايا المصابة بفايروس موزائيك التبغ واعتقد أنها بروتوزوا.
(4) فرضية السم الراشح والتي وضعها "بايرنك" سنة 1899 مفترضا أن المسبب هو مادة سامة غير خلوية ذائبة في الماء تعبر المرشحات البكتيرية أطلق عليها مصطلح "السائل الحي المعادي" Contagium vivum fluidum او "السم الراشح" Filterable virus.
(5) الفرضية الفسلجية والتي ضعها "داجر و كارير" Duggar & Karrer سنة 1921 مفترضين أن المسبب هو مادة كيميائية ايضية نباتية ناتجة عن اختلال فسلجي في خلايا النبات المريضة.
نشوء الفايروسات
أثار أصل الفايروسات وتطورها النشوئي Evolutionary development (يستعمل أيضا مصطلح Phylogeny) جدلا واسعا لدى الباحثين وذلك لعدم وجود أدلة مادية ملموسة عنها وهي الأحافير أو المتحجرات Fossils كما هو الحال مع الكائنات الأخرى حيث أن الاحافير هي السجلات التاريخية المحفوظة في الارض لأجساد الكائنات والتي تدل على كيفية تطور الكائنات الحية، ولازال أصل الفايروسات لغزا أمام الباحثين لذلك وضعت عدة فرضيات تفسر نشوء وتطور الفايروسات إلا أن كل هذه الفرضيات هي تكهنية غير مستندة على أدلة علمية دامغة بل على ملاحظات في التشابه التركيبي لمكونات الفايروسات مع بعض المكونات الخلوية والتماثل النسبي في اليات التضاعف والنشاط الحيوي للفايروسات مع بعض الأليات الخلوية، وفيما يلي وصف لهذه الفرضيات الثلاثة.
1. فرضية استقلالية الفايروسات
تفترض هذه الفرضية بان الفايروسات ظهرت كجزيئات رايبية RNA molecules وجدت قبل نشوء الخلايا على سطح الارض اي في فترة العالم قبل الخلوي Precellular world، حيث يعتقد انه قبل ظهور الخلايا البدائية كانت هناك مرحلة حدث فيها تطور للمواد غير الحية إلى جزيئات عضوية وبروتينات وأحماض نووية رايبية وهذا يعني ظهور الجزيئات الرايبية قبل الخلايا الأولية، وامتلكت هذه الجزيئات قابلية التضاعف الذاتي Self-replication، واقترحت هذه الفرضية أن الحياة نشأت وتطورت في بيئة معدنية توفرت فيها ظروف خاصة من جو دافئ ورطوبة مهدت لنشوء الحياة وبذلك ظهر الحامض النووي الرايبي RNA السلفي للفايروسات والذي تطورت منه أشكال الحياة وهكذا ووفق هذه الفرضية فان الفايروسات نشأت من تجميع العناصر أولية معدنية ذاتية التضاعف وربما نشأت الخلايا الأولية بنفس الطريقة أو تطورت من الفايروسات، (الشكل 1) كما تفترض هذه الفرضية أن الكائنات الحية لم تنبثق من البكتريا الأولية Primitive bacterium ولكن من "كائن أولي غير خلوي سلفي مشترك" سمي Last Universal Common Ancestor LUCA والذي انفصل عنه الفايروس الأولي، (الشكل 2) إلا أن هذه الفرضية يرفضها أغلب المهتمين بنشوء وتطور الحياة فهي ضعيفة الحجة وذلك لأن كل الفايروسات هي متطفلات اجبارية على الخلايا الحية مما يعني حتمية وجود الخلايا قبل الفايروسات.
ويبين الشكل (3) أن الخلايا حقيقية النواة قد ظهرت على الأرض في مرحلة متأخرة بعيدة عن مرحلة ظهور الخلايا بدائية النواة المتمثلة بالبكتيريا والاراكيا Arachaea، لذا فان التدرج الزمني التطوري Evolutionary timescale للفايروسات المتطفلة على الخلايا حقيقية النواة هو احدث بكثير من مثيله للفايروسات المتخصصة على الخلايا بدائية النواة.
أما الفايروسات المتطفلة على اللبائن الحديثة والطيور فيفترض أنها انحدرت من تلك التي كانت متطفلة على الداينوصورات، وبذلك فان هذه الفرضية تفترض أن الفايروسات هي كائنات طفيلية مستقلة لم تتطور من كائنات عضوية متطفلة لذا فهي تمثل مرحلة هامة من مراحل تطور المادة الحية حيث يعتقد انه في بواكير نشوء الحياة على الأرض وقبل إيجاد الكائنات الخلوية ظهر أولا الحامض النووي الرايبي RNA والذي طور بعد ذلك النشاطات الإنزيمية الرايبية Ribozymes ثم امتلك قدرة التضاعف الذاتي، وبذلك كانت الفايروسات هي أول الكائنات التي وضعت على عتبة الحياة بشكل جزيئات رايبية والتي تمكنت من التطفل حال ظهور الخلايا ثم اكتسبت جينات غطاؤها البروتيني وبذلك ظهر أول فايروس نباتي بقدرة الخالق الباري المصور، إن إحدى المفاهيم التي اعتمدت عليها هذه الفرضية هي الصلة الوثيقة القائمة بين الحامض النووي الفايروسي وجينوم العائل حيث يبدوا وكان خلايا العائل تفضل أن يتحكم بها الجينوم الفايروسي بدلا من أن تخضع لأحماضها النووية. إن الإجابة المعتادة عند الباحثين المختصين بالتطور عن السؤال " من أين جاءت الفايروسات ؟ هو : لا نعرف !.
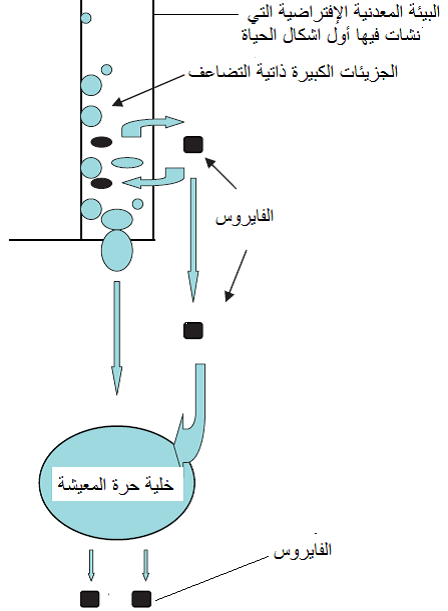
شكل (1): فرضية نشوء الفايروسات والخلايا من البيئة المعدنية ذاتية التضاعف، ثم تطفلها في مرحلة لاحقة على الخلايا حرة المعيشة.
الشكل مقتبس من Roossinck (2008).
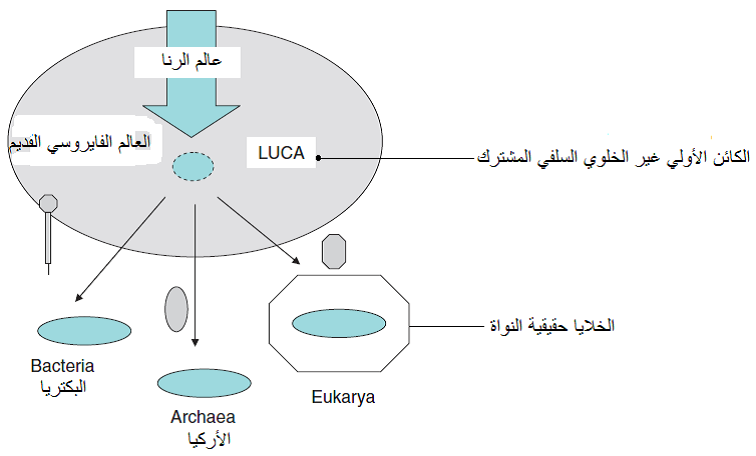
شكل (2): فرضية نشوء الحياة من الحامض النووي الرايبي، ثم نشوء الفايروسات الاولية منه، وكذلك نشوء الكائن الاولي غير الخلوي السلفي (LUCA) الذي نشأت منه الكائنات الخلوية الاولية.
الشكل مقتبس من Roossinck (2008).
2. الفرضية التراجعية أو الاختزالية
هي الفرضية التراجعية Reduction hypothesis التي تفترض ان الفايروسات كانت كائنات خلوية عضوية طفيلية ثم تراجعت نتيجة تطفلها المطلق على العائل لتتخلى عن معظم تراكيبها وتحتفظ فقط بالحامض النووي الذي يحمل صفاتها الوراثية مع غطاء يحميه، أي أنها نشأت بتطور تراجعي اختزالي Reductive evolution وبذلك نشأت الفايروسات البكتيرية من جينومات الخلايا بدائية النواة Procaryotes ونشات بقية الفايروسات ومنها الفايروسات النباتية من جينومات الخلايا حقيقية النواة Eucaryotes، (الشكل 3).
واعتمدت هذه النظرية على مبدأ أن التطفل الداخلي الخلوي Intracellular parasitism للفايروسات في خلايا عوائلها هي ظاهرة شائعة في متطفلات أخرى وهي الفايتوبلازما والركتسيا وليست صفة خاصة بالفايروسات وحكرا عليها ولكن يبدوا أن الفايروسات واصلت اعتمادها على الكائنات العائلة ففقدت معظم تراكيبها واحتفظت فقط بمادتها الوراثية، كذلك استندت هذه الفرضية على ندرة وجود الفايروسات المتطفلة على النباتات عارية البذور والنباتات البدائية مما يوحي بان الفايروسات النباتية ظهرت مع ظهور النباتات الأحدث وهي النباتات مغطاة البذور على الكرة الأرضية أي قبل 100 – 140 مليون سنة، كذلك فان حركة القارات في العصور الجيولوجية السحيقة ربما ساعدت على توزيع الأنواع الفايروسية وناقلاتها، وبذلك فإن هذه الفرضية قد انطلقت من الفكرة التي تقول بان الفايروسات هي متطفلات على الخلايا عليه فان المنطق يقول بعدم وجود أي فايروس قبل نشوء الخلايا على سطح الأرض علما بان هناك أدلة على وجود الخلايا الحية على الأرض قبل ما يقرب من 4 بليون سنة، (الشكل 4) عليه فهي تفترض أن أسلاف الفايروسات هي مكونات الخلايا والتي تمثل الأسلاف الجزيئية Molecular precursors للفايروسات الحالية، أو أنها انحدرت من الكائنات المجهرية المتطفلة داخل الخلايا.
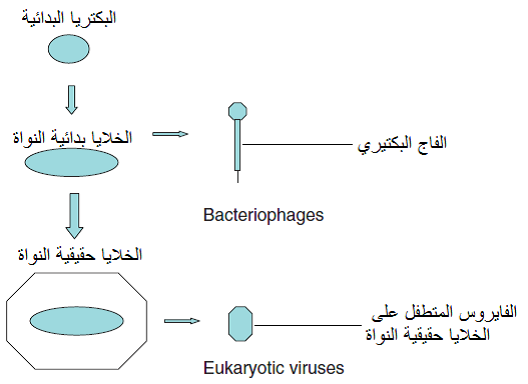
الشكل (3): نشوء الفايروسات النباتية من الخلايا الاولية حقيقية النواة ونشوء الفايروسات البكتيرية (الفاجات) من الخلايا بدائية النواة.
الشكل مقتبس من Roossinck (2008).

الشكل (4): المدى الزمني Timeline لتاريخ الارض والذي يبين التواريخ التقريبية المقدرة ببلايين السنين لنشوء الكائنات الحية.
الشكل مقتبس من Roossinck (2008).
3. فرضية التطور من العضيوات والمكونات الخلوية
يطلق عليها أيضا فرضية الهروب Escape hypothesis والتي تفترض أن أسلاف الفايروسات قد نشأت من أجزاء لمواد وراثية خلوية هربت من سيطرة الخلية حيث تشظت جينومات الخلايا السلفية البدائية الرايبية وكونت كروموسومات شبه مستقلة Semiautonomous Chromosomes والتي بدأت بالتضاعف بشكل مستقل لتنتقل بين الخلايا ولقد أنتج تكاثر الخلايا السلفية البدائية الرايبية في بداية نشوء الحياة مزيجا من الذرية بعضها يمتلك نظامي الكروموسومات والرايبوسومات والتي تطورت إلى الخلايا الأولية، والاخرى تمتلك إما الكروموسومات أو الرايبوسومات، والاخيرة لم تستمر بالحياة فيما تمكنت تلك التي تمتلك الكروموسومات على البقاء نتيجة تطفلها على الخلايا وهي أسلاف الفايروسات، وقد تكون أسلاف الفايروسات هي العضيوات الداخلية للخلايا أي الأسلاف الجزيئية والتي تمتاز باستقلاليتها النسبية عن الخلية لأنها تمكنت من تطوير قابليتها في التضاعف الذاتي بشكل مستقل عن سيطرة الخلية وبالتالي أصبحت طفيليات على تلك الخلايا وتوجد أدلة قوية على أن المايتوكوندريا والكلوروبلاستات في الخلايا حقيقية النواة هي منبثقة من الخلايا بدائية النواة ثم أصبحت عضيوات Organelles مستقلة داخل الخلايا حيث كانت أصلا متطفلة عليها ثم أصبحت متعايشة معها ثم فقدت قدرتها على انجاز العديد من وظائفها الحيوية وفقدت جيناتها التي تشفر معظم بروتيناتها التي استغنت عنها وبالتالي تخلت عن آلية تخليق البروتين وتحولت إلى فايروسات، كذلك تفترض ان الأسلاف المرشحة للفايروسات هي الجزيئات الرايبية المراسلة mRNA وجزيئات البلازمد Plasmids والترانسبوسونات Transposones وهي جزيئات الدنا DNA المستقلة (الشكل 5) إذ يلاحظ أن جينومات العديد من الفايروسات الرايبية موجبة التوجهssRNA (+) المتطفلة على الخلايا حقيقية النواة تمتلك خصائص الأحماض الرايبية المراسلة لتلك الخلايا، فهناك تماثل في النهاية 5 لجينوم فايروسات Tobamoviruses و Reoviruses مع الأحماض الرايبية المراسلة للخلايا التي تتطفل عليها حيث أن كلاهما يمتلك نهاية مغطاة وعديدة الادينيل Capped end and Polyadenylated وبذلك فهي تماثل تركيب الأحماض المراسلة لخلايا عوائلها، وهناك اعتقاد بان الفايروسات ذات جينوم الدنا قد انحدرت من البلازمد والذي يطلق عليه أيضا اسم "الربلكون" Replicon" أو يسمى "العامل الكروموسومي الإضافي" Extrachromosomal factor وهو جزيئة الدنا الحلقية التي توجد في أنواع من الخلايا حقيقية أو بدائية النواة ويتراوح حجمها بين ا– 200 كيلو قاعدة (Kilo base (Kb ووجد أن البلازميدات البكتيرية تحمل الجينات المتخصصة بإنتاج الزوائد البروتينية المتخصصة Sex pilus التي تربط كل خليتين بكتيريتين عند التزاوج مما يسمح بعبور البلازمد من الخلية الواهبة إلى الخلية المستقبلة لذلك هناك رأي يقول بان الفاجات البكتيرية الخيطية Tailed phages ربما انحدرت من البلازميدات أو من الزوائد البروتينية الجنسية، ولكن بالمقابل هناك رأي يقول بان الزوائد الجنسية هي التي انحدرت من الفاجات.
أما الترانسبوسونات TranspoSons فهي تعاقبات لجزيئة دنا متحركة موجودة في جينومات الخلايا حقيقية وبدائية النواة ولها قدرة الحركة من جزء من الجينوم إلى جزء آخر بالية القطع واللصق Cut & Past mechanism، وربما تطورت الفايروسات بهذه الطريقة حيث اكتسبت خلال تطورها بعض جينات جينوم العائل وبضمنها جينات الكابسيد وإنزيم البوليميريز، (الشكل 5) أما السلف الأخر المحتمل جدا لنشوء الفايروسات فهي الترانسبوسونات العكسية Retrotransposons وهي تعاقبات شبيهة بفايروسات الاستنساخ العكسي Retrovirus-like Sequence موجودة في جينومات الخلايا حقيقية النواة وتتكون من جينات تشفر إنزيم الاستنساخ العكسي Transcriptase وإنزيم التكامل Integrase لذلك يعتقد أنها أسلاف فايروسات الاستنساخ العكسي، علما بان الترانسبوسونات العكسية توجد في الجينومات البشرية والحيوانية واللافقارية وفي النباتات والفطريات.
يتبين مما سبق أن هناك ثلاث مسارات محتملة لنشوء وتطور الفايروسات وهي
(1) أنها جزيئات رايبية RNA وجدت قبل نشوء الخلايا على سطح الأرض
(2) تطورت من كائنات مجهرية دقيقة
(3) تطورت من مكونات خلوية.
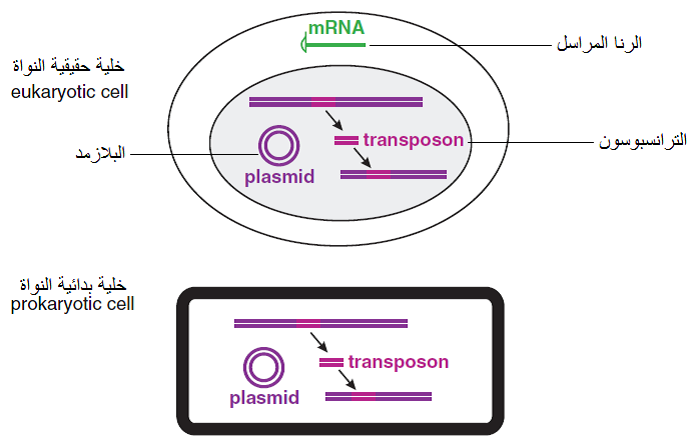
الشكل (5): المكونات الخلوية الجينية في الخلايا حقيقية النواة وبدائية النواة المتوقعة ان تكون اسلافا للفايروسات.
الشكل مقتبس من Carter و Saunders (2007).
اليات تطور الفايروسات بعد النشوء
تخضع الفايروسات كغيرها من الكائنات الحية إلى حتمية التغير الوراثي والتطور لتستطيع التكيف مع البيئات الجديدة، لذا فان التغيير الذي حصل للفايروسات بعد نشوئها ربما حصل وفق إحدى المسارات الأتية:
1. التطور بالتطفير
تحصل الطفرات Mutations بسبب تغير يحصل عند تضاعف الحامض النووي مما يسبب ظهور أفراد مغايرة للإباء ثم يعقب ذلك حصول الانتخاب الطبيعي بين تلك الافراد الجديدة Natural Selection، وبذلك فان الطفرة الأصلح هي التي ستبقى وتظهر حالة التلاؤم Fitness الأفضل مع البيئة.
2. التطور بإعادة الارتباط الجيني بين سلالات فايروسيه
إعادة الارتباط الجيني Recombination هي عملية ارتباط قطع من جينوم سلالتين فايروسيتين مختلفتين تؤدي إلى إنتاج جينوم جديد من الجينومين الأبويين وهذا يحصل في الفايروسات عند حدوث الإصابة الخليطة للنبات بسلالتين لنفس النوع الفایروسي معا ويسمى الفايروس الجديد المنتج "الفايروس المُشكل" Recombinant Virus.
3. التطور بإعادة التشكيل الجينومي بين سلالات فايروسيه
إعادة التشكيل Reassortment هي نوع خاص من آلية إعادة الارتباط أعلاه والتي تحصل في الفايروسات ذات الجينوم المقسم الذي تكون كل قطعه الجينومية معبأة في جسيمة فريون واحدة كما في فايروسات Reoviruses و Bunyaviruses، فإذا ما أصيبت الخلية بوقت واحد بسلالتين لفايروس معين فان الفريونات الوليدة (الذرية) قد تحوي على مزيج من قطع جينومية هجينة اكتسبتها من الجينومين الأبويين، ويسمى الجينوم الجديد "المتشكل" Reassortant genome.
4. التطور باكتساب جينات خلوية
أدى اكتشاف التماثل بين بعض البروتينات الفايروسية والخلوية إلى هذا الاستنتاج وهو قيام الفايروسات باصطياد جينات العائل، وهذا ما حصل مع بعض الفايروسات البشرية وهي فايروسات الاستنساخ العكسي Retroviruses إلا انه لم يلاحظ ذلك لحد الأن مع الفايروسات النباتية.
أدت آليات التطوير الوراثي التي ذكرت أعلاه إلى نشوء العديد من الفايروسات الجديدة التي لم تكن موجودة سابقا وذلك نتيجة لإصابة أنواع جديدة من العوائل وهذا ما حصل مع بعض الفايروسات البشرية مثل فايروس نقص المناعة 2-HIV و 1-HV وهو غير مستبعد حصوله مع الفايروسات النباتية حيث تتوفر العديد من الأدلة على أن فايروس الذبول المبقع للطماطة (TSWV) هو أحد الفايروسات الناشئة والمتطورة حديثا خاصة وأنه أصبح أحد الفايروسات الخطرة وبائيا على العديد من المحاصيل الاقتصادية في السنوات الأخيرة.
الشجرة الوراثية للفايروسات
إن دراسة تطور الفايروسات وفهمه يتطلب دراسة التعاقب الجينومي للفايروسات ووضع الشجرة الوراثية (الشجرة التطورية) Phylogenetic tree التي تبين درجة القرابة بين الفايروسات، ويبين الشكل (6) النوعين المعروفين منها وهما:
(1) الشجرة الوراثية الجذرية Rooted Phylogenetic tree وهي التي تبدأ بجذر يمثل "سلف" Ancestor الفايروسات الموجودة في تلك الشجرة.
(2) الشجرة الوراثية غير الجذرية Unrooted Phylogenetic tree وفي هذا النوع لا يوجد سلف معين للفايروسات في الشجرة، وتشير أفرع الشجرة التطورية إلى صلة القرابة بين الفايروسات في التعاقبات النيوكليوتايدية وهذه الأفرع قد تكون مدرجة Scaled branchs حيث تمثل أطوال تلك الأفرع المسافات الوراثية Genetic distances بين التعاقبات النيوكليوتيدية، أو تكون الأفرع غير مدرجة ,Unscaled branches اما الارقام التي تظهر في الشجرة الوراثية فتشير إلى النسبة المئوية لتطابق صفة التشخيص المختارة بين الفايروسات.
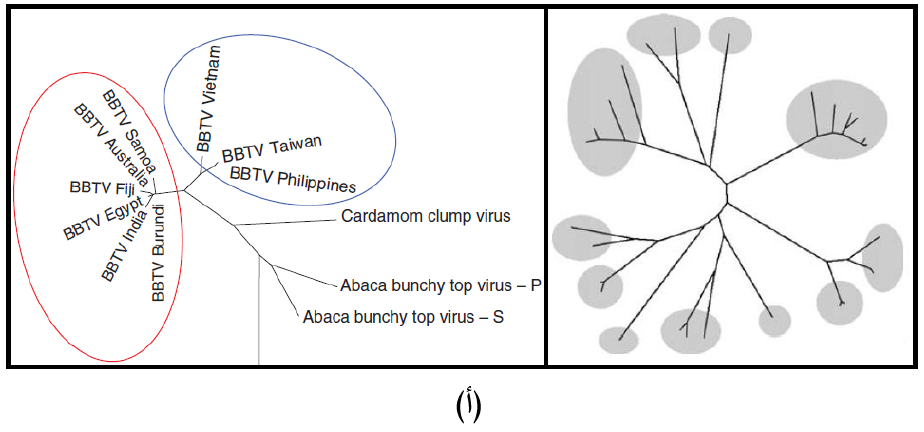
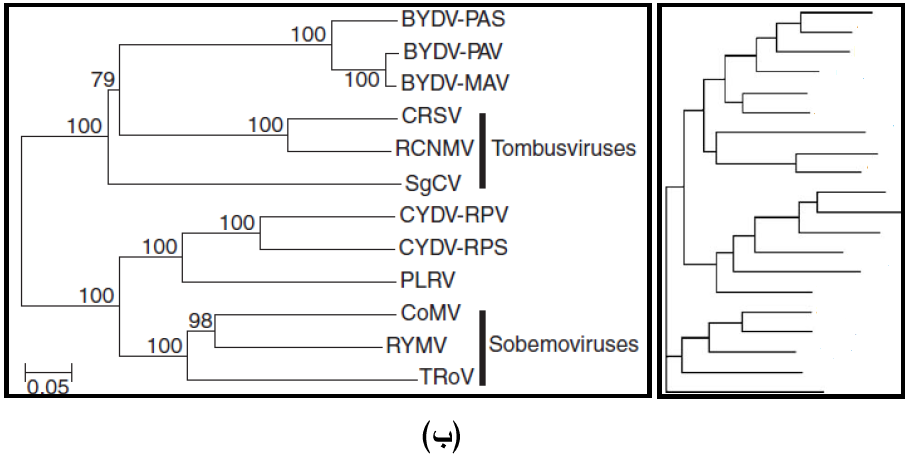
الشكل (6): الشجرة الوراثية (أ) يظهر الشكل الايمن مخطط عام للشجرة الوراثية الجذرية فيما يظهر الشكل الايسر مثالا للعلاقة الوراثية الذي تظهره هذه الشجرة بين سلالات فايروس تكتل قمة الموز (BBTV) وانواع الفايروسات المبينة في الشكل (ب) يظهر الشكل الايمن مخطط عام للشجرة الوراثية غير الجذرية فيما يظهر الشكل الايسر مثالا للعلاقة الوراثية الذي تظهره هذه الشجرة بين فايروسات Tombusviruses و Sobemoviruses.
الشكل مقتبس من Mahy و Van Regenmortels (2008).
تعريف الفايروس النباتي
وضعت العديد من التعاريف للفايروس النباتي، إلا أن أشملها هو التعريف الذي وضعه Matthews (1981)وهو "الفايروس كائن طفيلي مؤلف من جزيئة واحدة أو عدة جزيئات جينومية متجانسة ومتكاملة حاملة للجينات، وهي إما حامض نووي رايبي مفرد أو مزدوج الخيط ssRNA or dsRNA أو حامض نووي منقوص الأكسجين مفرد أو مزدوج الخيط ssDNA or dsDNA، والتي يضمها غطاء بروتيني واقي Coat protein يسمى "الكابسيد" Capsid وتمتلك الفايروسات المعقدة غلاف إضافي Envelope مكون من البروتين والليبيد (الليبوبروتين) يغلف الكابسيد، وان هذه الجزيئات الجينومية قادرة على انجاز تضاعفها الذاتي داخل الخلايا الحية فقط بالاعتماد على الية تخليق البروتين في خلايا العائل Host protein synthesizing machinery فهي لا تمتلك الآلية الجزيئية الذاتية للتضاعف، كما تحصل على مكوناتها الأساسية وهي النيوكليوتايدات والأحماض الأمينية من خزين الخلية وعليه فإن الفايروسات تتكون من وحدات يتم تجميعها وذلك بعد تخليقها من قبل خلايا العائل، أما بقية الكائنات المتطفلة فتتألف أجسامها من وحدات بنائية تخلقها بنظامها الايضي الخاص بها، لا تمتلك الفايروسات النباتية نظام خزن وإطلاق الطاقة المتمثل بمركب الادينوسين ثلاثي الفوسفات ATP , Adenosine triphosphate، وتمتلك الفايروسات قدرة الانتقال من النباتات المصابة بها وإحداث العدوى في أفراد جديدة.
تسمى الجسيمة الفايروسية الكاملة "الفريون" Virion ويعرف بأنه "الفايروس القادر على إحداث الإصابة والكامل تركيبيا أي يحوي على الحامض النووي والغطاء البروتيني (الكابسيد) في الفايروسات البسيطة، والحامض النووي والكابسيد والغلاف الليبوبروتيني في الفايروسات المغلفةEnveloped viruses. .
إن الفايروسات النباتية هي إحدى اصغر الكائنات المتطفلة على النباتات رغم وجود قلة من الفايروسات التي يزيد طولها عن أنواع الركتسيا والفايتوبلازما الممرضة للنبات ومنها فايروس ترستيزا الحمضيات (CTV) وهو أطول فايروس نباتي مكتشف حيث يبلغ طوله بحدود 2000 نانومتر لذلك فهو الوحيد من الفايروسات النباتية الذي يمكن رؤيته بالمجهر الضوئي. ولتخيل حجم الفايروسات النباتية المتناهي في الصغر فإن نسبة حجمها إلى حجم بالون صغير كنسبة حجم البالون إلى الكرة الأرضية، علما بان الأخيرة أكبر من البالون بمقدار 10 مليون ضعف، (الشكل 7)، ويبين الشكلين (8 و 9) مقارنة بين حجوم جينومات الفايروسات مع حجوم جينومات الكائنات الحية محسوبة بعدد نيوكليوتايدات الجينوم وكذلك حجوم مجموعة مختارة من فايروسات النبات مقارنة مع حجوم أهم ممرضات النبات داخل الخلية النباتية.

الشكل (7): مقارنة حجمية بين جسيمة فايروسية وبالون صغير والكرة الارضية علما ان نسبة حجم البالون الى حجم الفايروس هي 710 اي انه اكبر من الفايروس بمقدار 10 مليون مرة، والارض اكبر من البالون ايضا بمقدار 10 مليون مرة.
الشكل مقتبس من Carter و Saunders (2007).

الشكل (8): مقارنة بين حجوم جينومات الفايروسات مع حجوم جينومات البكتريا والفطريات ومعظم النباتات والحيوانات بدلالة اعداد النيوكليوتايدات.
الشكل مقتبس من Hull (2002).

الشكل (9): مقارنة بين حجوم عدة انواع من الفايروسات مع بعض اهم ممرضات النبات داخل خلية النبات العائل.
الشكل مقتبس من Agrios (2005).



|
|
|
|
التوتر والسرطان.. علماء يحذرون من "صلة خطيرة"
|
|
|
|
|
|
|
مرآة السيارة: مدى دقة عكسها للصورة الصحيحة
|
|
|
|
|
|
|
نحو شراكة وطنية متكاملة.. الأمين العام للعتبة الحسينية يبحث مع وكيل وزارة الخارجية آفاق التعاون المؤسسي
|
|
|