
آخر المواضيع المضافة

 النبات
الحيوان
الأحياء المجهرية
علم الأمراض
التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحياتية النانوية
علم الأجنة
الأحياء الجزيئي
علم وظائف الأعضاء
الغدد
المضادات الحيوية
النبات
الحيوان
الأحياء المجهرية
علم الأمراض
التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحياتية النانوية
علم الأجنة
الأحياء الجزيئي
علم وظائف الأعضاء
الغدد
المضادات الحيوية| الارتباط والتعابر والخرائط الوراثية Linkage, Crossing Over and Genetic Maps |


|
|
أقرأ أيضاً
التاريخ: 19-10-2020
التاريخ: 25-11-2015
التاريخ: 11-11-2015
التاريخ: 21-3-2018
|
الارتباط والتعابر والخرائط الوراثية Linkage, Crossing Over and Genetic Maps
وجد بونت Punnett وباتسون Bateson عام 1906م بأن زوجين مختلفين من الاليلات لا تنعزل بصورة مستقلة في البزاليا الحلوة , أي أنها لا تخضع لمبدأ الانعزال الحر لمندل . وكذلك أوضحت الدراسات على ذبابة الفاكهة من قبل موركان Morgan والعاملين معه للفترة 1910-1915م بأنه لا يمكن تطبيق مبدأ الانعزال الحر كلياً بسبب وجود بعض التحديدات لبعض التضريبات لزوجين أو أكثر من الجينات . وفي هذه التضريبات كان هناك دليل واضح للتراكيب الأبوية لتبقى مرتبطة بنسبة عالية ولكنها تنتج تراكيب جديدة بنسبة واطئة . وأسست جماعة موركان نظرية الارتباط Linkage والتعابر Crossing over على أسس خلوية ثابتة ومباشرة بعد تأسيس نظرية الارتباط والتعابر في ذبابة الفاكهة , وجد إن كائنات حية أخرى تتبع نفس النظرية . ودعمت هذه الدراسات نظرية الكروموسوم الوراثية وأدت إلى إعداد الخرائط الوراثية التي صورت العلاقات بين الجينات في الكروموسومات . وأول ما عُرف الارتباط بأنه ميل الجينات الغير أليلية الواقعة على نفس الكروموسوم أو في نفس زمرة الارتباط الدخول معاً (أي بتراكيب أبوية) بنسبة أعلى مما يتوقع من الانعزال الحر . تكون الجينات الغير أليلية مرتبطة بسبب وقوعها على نفس الكروموسوم ولذا تحاول أ تبقى معاً أثناء الانقسام الميوزي وتدخل نفس المشيج . وتكون دراسة الارتباط سهلة وبسيطة في حالة بقاء الجينات معاص على الدوام وعلى نفس الكروموسوم وبذا يحدث الارتباط التام Complete linkage . ولكن لا يحدث ذلك اعتيادياً , بل يحدث تبادل أو تعابر Crossing over بين أزواج الكروموسومات المماثلة في أكثر النباتات والحيوانات وبذا يحدث الارتباط غير التام Incomplete linkage وقد أصبح من المعلوم الآن أن جينات الكائن الحي تتوزع على عدد كروموسوماته وسميت مجموعة الجينات المرتبطة في كروموسوم واحد المجموعة (الزمرة) الارتباطية (Linkage group) ولذا فإن عدد المجاميع الارتباطية لحشرة ذبابة الفاكهة يساوي أربعة وللإنسان يساوي 23 وهكذا .
الارتباط التام Complete Linkage
يعتبر الارتباط تاماً عندما تكون الجينات متقاربة جداً وتنتقل معاً على الدوام من جيل لآخر . فمثلاً كل طفرات الكروموسوم الرابع في ذبابة الفاكهة من نوع D. melanogaster تنعزل قليلاً أو لا تنعزل كل واحدة عن الأخرى أثناء الانتقال . فعند تضريب ذبابة تحمل الجينات التي تنتج أجنحة منحنية وشعيرات محلوقة الواقعة على الكروموسوم الرابع ونرمز لها بـ b s/ b s مع ذبابة طبيعية , سينتج F1 ذو مظهر طبيعي ومتباين الزيجة للجينات الطافرة . وعندما ضرب هذا الـ F1 مع سلالة منحنية ومحلوقة بحالة متماثلة الزيجة كانت الذرية ذات نمط ظاهري إما منحنية ومحلوقة أو طبيعية لكلا الصفتين وعملياً لو تظهر ذبابات ذات أجنحة منحنية وشعيرات طبيعية أو ذات أجنحة طبيعية وشعيرات محلوقة . أن عدم وجود أي درجة من الانعزال الحر بين هذين الزوجين من الجينات يكون دليلاً على الارتباط الشديد بينهما . ويوجد هذا الارتباط عندما يكون جين المنحنية وجين المحلوقة مشتركان معاً على نفس الكروموسومات , وكذلك عندما يكون كل منهما مشتركاً مع أحد الجينين الطبيعيين على نفس الكروموسوم , كما في المثال التالي : يمكن إجراء تضريب خلفي لـ F1 (الناتج من تضريب ذبابات متماثلة الزيجة ذات أجنحة منحنية وشعيرات طبيعية بذبابات ذات أجنحة طبيعية وشعيرات محلوقة) مع ذبابات من السلالة المنحنية والمحلوقة . وتكون ذرية هذا التضريب الخلفي ذات نمط ظاهري مشابه إلى الأبوين الأصليين : أي ذات أجنحة منحنية وشعيرات طبيعية وأخرى ذات أجنحة طبيعية وشعيرات محلوقة.
كذلك لوحظ الارتباط التام الناتج من عدم وجود العبور لكل أزواج الجينات في كل من زمر الارتباط الأخرى عندما تستعمل ذكور تستعمل ذكور ذبابة الفاكهة كأب بحالة متباينة الزيجة , فمثلاً يقع جين العيون الأرجوانية وجين الأجنحة الأثرية في زمرة الارتباط الثانية وهما متنحيان إلى جين العيون الحمراء وجين الأجنحة الطويلة الطبيعيتين . فمن تضريب ذبابة متماثلة الزيجة للأرجوانية والأثرية على ذبابة طبيعية (العيون الحمراء والأجنحة الطويلة) , ينتج كل ذرية F1 متباينة الزيجة وطبيعية . ومن التضريب الخلفي لذكر F1 على أنثى متماثلة الزيجة للأرجوانية والأثرية , ينتج نمطان ظاهريان فقط : أرجوانية وأثرية (متماثلة الزيجة) وطبيعية للصفتين (متباينة الزيجة) .
ويظهر الارتباط التام كخاصية في ذكور ذبابة الفاكهة وفي الجنس المتباين المشيج Heterogametic sex لأنواع قليلة أخرى (كإناث دودة الحرير) .
الارتباط غير التام Incomplete Linkage
أتضح لدينا بأن الارتباط التام بين الجينات على نفس الكروموسوم نادر في معظم الأنواع التي تتكاثر جنسياً . وكقاعدة يكون الارتباط غير تام , ويكون الانعزال الحر جزئياً بين أزواج الجينات في أكثر زمر الارتباط , وكان موركان أول من لاحظ ذلك بوضوح في التضريبات بين السلالة ذات العيون البيضاء والأجنحة المصغرة والسلالة الطبيعية في ذبابة الفاكهة . إن هذه الصفات مرتبطة بالجنس . وعندما أزواج موركان ذكر ذو عيون بيضاء وأجنحة مصغرة على أنثى طبيعية وذات النمط البري كانت ذكور وإناث F1 من النمط البري . فإذا كان الانعزال بين جين البيضاء وجين المصغرة على كروموسوم X انعزالاً حراً , فيجب أن تظهر ذكور F2 بأربعة أنماط ظاهرية : نمط بري w+ m+ , بيضاء ومصغرة w m , بيضاء w m+ , مصغرة w+ m وبتردد متساوي . أما إذا كان الارتباط بين جين البيضاء وجين المصغرة ارتباط تاماً , فيجب أن تظهر الأنماط الأبوية الأصلية فقط بين ذكور F2 . وبالحقيقة لاحظ موركان بأن نتائج F2 تقع بين الارتباط التام والانعزال الحر لهذين الزوجين من الجينات حيث نتج 1190م ذكر في F2 , كان منها 37,6٪ بيضاء أو مصغرة والتي تمثل الأنماط الأبوية .
أفترض موركان بأن الجينات توجد بترتيب خطي على طول الكروموسوم ، وعليه يكون الارتباط علاقة فيزياوية بين الجينات التي يمكن تحويرها بواسطة التعابر الفيزيائي بين أزواج الجينات على الكروموسومات المتماثلة ، ونظرية التعابر هذه تتفق جيداً مع النظرية التي قدمت من قبل جانسن Janssen في عام 1909م والتي تنص على إن التصالبات Chiasmata الملحوظة خلوياً تمثل نقاط تعابر بين الكروموسومات المتماثلة .
الأساس الخلوي للتعابر Cytological Basis Of Crossing Over
قدم الدليل الوراثي للارتباط والتعابر قبل توضيح الأساس الخلوي لهما . حيث أفترض بأن الجينات المرتبطة تترتب ترتيباً خطياً على كروموسوماتها الخاصة وأعتبر التعابر الوراثي نتيجة لتبادل أجزاء بين الكروموسومات المتماثلة . وفي البداية لم يمكن برهان هذه النظرية خلوياً لأن الكروموسومات المتماثلة تظهر عند فحصها مجهرياً بأنها متشابهة تماماً , وكان مستحيلاً ملاحظة تبدل محلات أجزاء الكروموسومات لحين دمج علامات مرئية بالمجهر على الكروموسومات . وأنجز ستيرن Stern عام 1931م البرهان الخلوي للتعابر بتجاربه الكلاسيكية على ذبابة الفاكهة , وكذلك أنجز كريكتون Creighton وماك كلينتوك Mc Clintock عام 1931م نفس الشيء بتجاربهما على الذرة الصفراء .
حصل ستيرن على ذبابات ذات كروموسومات X التي يمكن تمييزها بالمجهر حيث أن جزءاً من كروموسوم Y قد أنكسر ثم التحق بأحد كروموسومي X في السلالة الأولى . وكسر جزء من كروموسوم X بمعاملته بالأشعة السينية ثم ألحق بالكروموسوم الرابع الصغير في السلالة الثانية . ومن تضريب هاتين السلالتين ينتج ذبابات الجيل الأول التي يمكن تمييز كل من كروموسومي X للإناث بالفحص المجهري , وبذا أصبح من الممكن برهان حدوث التعابر خلوياً . حيث أخذ ستيرن إناثاً متباينة الزيجة إلى الجين c الذي ينتج العيون القرنفلية Carnation والجين B الذي ينتج العيون القضيبية Bar , يرتبط هذان الجينان بكروموسوم X . ضُربت هذه الإناث ذات العيون الحمراء اللون والقضيبية الشكل على الذكور ذات العيون القرنفلية اللون والطبيعية الشكل . درست الأنماط الظاهرية للذبابات الناتجة من هذا التضريب وعزلت الذبابات الحاوية على تراكيب جديدة الناتجة من التعابر , وعند فحص خلايا هذه الذبابات بالمجهر أظهرت تبدل في كروموسومي X ناتج من تبادل بين الكروموسومات المتماثلة أثناء الانقسام الميوزي وبذا قدم الأساس الفيزياوي أو الخلوي للتعابر .
أستعمل كريكتون وماك كلينتوك عقدة على نهاية كروموسوم معين وشدود مرئي على الكروموسوم المثيل له كعلامات خلوية في الذرة الصفراء . وعلى العموم كانت نتائجهم مشابهة لنتائج ستيرن .
الكشف عن الارتباط والتعابر Detection of Linkage and Crossing Over
يوجد طريقتان مناسبتان لتعيين الارتباط والتعابر في الكائنات الحية ثنائية المجموعة الكروموسومية :
الفائدة الرئيسية من طريقة دراسة معلومات تضريب الاختبار هو تجهيزها لنسب بسيطة , والتي تكون 1 : 1 : 1 : 1 في حالة حدوث التوزيع الحر لجينين مختلفين . فمثلاً يقع الجين vg للجناح الأثري في ذبابة الفاكهة من نوع D. melanogaster على الكروموسوم الثاني : بينما يقع الجين e للجسم الابنوزي اللون على الكروموسوم الثالث . ففي إحدى التجارب ضُربت أنثى طبيعية النمط الظاهري ومتماثلة الزيجة بذكر أثري الجناح و أبنوزي الجسم ثم ضُربت إناث F1 بذكور أثرية الأجنحة وأبنوزية الجسم المتنحيتين , ونتجت النسبة 1 : 1 : 1 : 1 وهي المتوقعة عندما يكون الجينان vg و e غير مرتبطين وبما أن النسبة 1 : 1 : 1 : 1 تكون خاصية التوزيع الحر , فالجينات الغير معروف موقعها وتعطي مثل هذه النتائج في تضريب الاختبار , يمكن الافتراض بأنها تقع على كروموسومات مختلفة . بينما يدل الانحراف المهم إحصائياً (المعين باختبار X2) عن النسبة 1 : 1 : 1 : 1 على شيء ما غير الانعزال الحر , ويكون الارتباط البديل الأكثر احتمالاً .
ويمكن توضيح البديل للانعزال الحر بالتجربة التالية : في ذبابة الفاكهة ضربت أنثى متماثلة الزيجة ذات أجنحة مستقيمة (نمط بري) وجسم رمادي (نمط بري) بذكر ذو أجنحة متجعدة Curled (وهي متنحية إلى الأجنحة المستقيمة ويرمز لها بـ cu) وجسم أبنوزي Ebony (وهو متنحي للجسم الرمادي ويرمز له بـ e) ونتجت إناث F1 التي ضربت بذكور ذات أجنحة متجمدة وجسم أبنوزي (أي تضريب اختبار) فنتجت نسبة مقاربة إلى :
أربعة مستقيمات ,رمادي : واحد مستقيمة , أبنوسي : واحد متجعدة , رمادي : أربعة متجعدة , أبنوزي . تدل هذه النتيجة على أن cu و e كانا على نفس الكروموسوم في F1 وبطور الازدواج Coupling phase .
وبتجربة أخرى كان التضريب بين إناثاً ذات أجنحة مستقيمة وأجساماً أبنوزية وذكوراً ذات أجنحة متجعدة وأجساماً رمادية .
ثم ضربت إناث F1 بذكور تحمل الصفتين المتنحيتين (وهذا هو تضريب اختبار أيضاً) .
أعطت نتائج تضريب الاختبار نسبة مقاربة إلى :
واحد مستقيمة ، رمادي : أربعة مستقيمة ، أبنوزي : أربعة متجعدة ، رمادي : واحد متجعدة أبنوزي كالمتوقع عندما يكون ترتيب الجينين بطور التنافر Repulsion phase في F1 . بعد معرفة إن جينين يقعان على نفس الكروموسوم ، يتم تقدير موقعهما النسبي بواسطة حساب تردد التعابر بينهما . وعند تحويل نسبة التعابر إلى موقعهما النسبي في الكروموسوم يكون نسبة واحدة من التراكيب الجديدة تساوي وحدة مسافة واحدة على خريطة الارتباط . ويكون عدد وحدات التعابر بين مكانين Two Loci لجينين مساوياً لنسبة الذرية التي تنتج من التبادل بين المكانين . فإذا حصلنا على أرقام مثالية ( 40 مستقيمة ، رمادي و 10 مستقيمة ، أبنوزي و 10 متجعدة ، رمادي و 40 متجعدة ، أبنوزي ) للتجربة السابقة بطور الازدواج فيمكن تمييز الأنماط الأبوية والتراكيب الجديدة كما يلي :
مستقيمة ، رمادي 40 أنماط أبوية Parental types
متجعدة ، رمادي 10 تراكيب جديدة Recombinations
مستقيمة ، أبنوزي 10 تراكيب جديدة Recombinations
متجعدة ، أبنوزي 40 أنماط أبوية Parental types
وعند دمج الأنماط الأبوية ينتج 80 بالمئة ، وعند دمج التراكيب الجديدة ينتج 20 بالمئة . وعليه تكون قوة الارتباط بين هذين المكانين 80 بالمئة والتراكيب الجديدة 20 بالمئة . وتكون نتائج تضريبات طور التنافر مشابهة إلى نتائج تضريبات طور الازدواج . استعملت أرقام مثالية في المثال المذكور أعلاه لتوضيح فكرة حساب الارتباط والتعابر إلا إن المعلومات (الأرقام) من التجارب الحقيقية لا تكون منتظمة مثل انتظام الأرقام المثالية . وأول معلومات حصل عليها باتسون وبونيت التي أوضحت الانحراف عن الانعزال الحر كانت من جيل F2 الناتج من تضريب ثنائي الهجين في البزاليا الحلوة . ومثل هذه المعلومات تكون صحيحة بنفس درجة صحة المعلومات من تضريب الاختبار لاكتشاف الارتباط وتقدير درجته ، ولكن دراستها أكثر إرباكاً بسبب النسبة 9 : 3 : 3 : 1 الأكثر تعقيداً من النسبة 1 : 1: 1: 1 التي تقارن النتائج معها . إلا إن معلومات F2 من تضريب ثنائي الهجين قد يحصل عليها بسهولة أكثر من معلومات تضريب الاختبار في بعض الأنواع وخاصة في النباتات ذات الإخصاب الذاتي كالحنطة والشعير ، حيث يكون إجراء تضريب الاختبار متعباً ومستهلكاً للوقت في هذه النباتات بإزالة المتوك والتلقيح باليد ، بينما يكون من السهل جداً السماح إلى نباتات F1 للإخصاب الذاتي لإنتاج نباتات F2 . ويوجد طريقتان لتقدير درجة الارتباط من معلومات F2 .
أ . طريقة الجذر التربيعي Square Root Method
يمكن استعمال تردد الأنماط الظاهرية المتنحية المزدوجة (مثل rr ss) في F2 لتقدير تردد الأمشاج التي لم يحصل فيها التعابر عندما يكون F1 في طور الازدواج الذي يساوي إلى
ثم يطرح العدد الناتج من واحد فينتج قيمة التعابر التي يمكن تحويلها إلى نسبة مئوية . ولتقدير تردد الأمشاج التي يحصل فيها التعابر عندما يكون F1 في طور التنافر الذي يساوي إلى  ثم يحول العدد الناتج إلى نسبة مئوية .
ثم يحول العدد الناتج إلى نسبة مئوية .
ب . طريقة نسبة الحاصل Product-ratio method
وبهذه الطريقة تستعمل جداول خاصة وطرق حسابية تتضمن تردد الأنماط الظاهرية الأربعة في F2 الناتجة من تضريب ثنائي الهجين وهي R-ss , R-S rr ss , rr S- , والتي يرمز لها بـ d , c , b , a على التوالي .
ولمعلومات الازدواج :
ولمعلومات التنافر :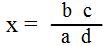
ويطلق على x نسبة الحاصل Product ratio وباستعمال جداول نسبة الحاصل يمكن استخراج ما يقابل قيمة x من نسبة التعابر أو التراكيب الجديدة .
تضريب الثلاثة نقاط Three-point cross
يمكن إجراء التضريب الثلاثة نقاط في حالة توفر ثلاثة جينات (c, b, a) التي تقع في نفس زمرة الارتباط لغرض تعيين المسافات بينهم ومواقعهم النسبية ورسم الخريطة الوراثية . يتوقع من التضريبات بين أفراد تحمل c+ , b+ , a+ بحالة متماثلة الزيجة مع أفراد تحمل c, b, a وبحالة متماثلة الزيجة إنتاج ذرية F1 بحالة متباينة الزيجة .
ويتوقع من تضريب الاختبار لـF1 بأفراد متنحية (c, b, a) ومتماثلة الزيجة أنتاج ذرية التي يمكن تصنيفها إلى أنماط أبوية وتراكيب جديدة . ولثلاثة جينات مرتبطة , تنحرف النتائج المتوقعة عن النسبة 1 : 1 : 1 : 1 : 1 : 1 : 1 : 1 الناتجة من تضريب ثلاثي الهجين وبافتراض التوزيع الحر . ففي أحدى تجارب ذبابة الفاكهة ضربت إناث التي تحمل ثلاثة جينات متنحية وبحالة متماثلة الزيجة وهي cu (الأجنحة المتجعدة Curled wings) وsr (الجسم المخطط Striped body) وe (الجسم الابنوزي Ebony body) بذكور تحمل الجينات السائدة وبحالة متماثلة الزيجة وهي :
cu+ (مستقيمة الأجنحة Straight wings) وsr+ (الجسم غير مخطط Not striped) وe+ (الجسم الرمادي Gray) . ثم اجري تضريب الاختبار لإناث F1 بذكور تحمل الصفات المتنحية : الأجنحة المتجعدة , الجسم المخطط , الجسم الابنوزي , ونتجت الأنماط الظاهرية والوراثية وبالشكل التالي :
وتتبع الخطوات الأساسية التالية لتحليل نتائج تضريب الثلاثة نقاط :

وتعكس هذه النسب وحدات المسافات بين الجينات على الكروموسوم اللازمة لإعداد خريطة الكروموسوم الوراثية . يمكن البدء بمنطقة sr cu شكل (1) حيث ينتج التعابر المفردSingle crossing over بين sr cu الصنوف 3 , 4 , 7 , 8 , ومجموع أعداد التراكيب الجديدة لهذه الصنوف وهو 207 (107 + 97 + 1 + 2 = 207) من المجموع الكلي البالغ 1926 ذبابة . وعليه تكون النسبة المئوية التعابر في المنطقة cu sr مساوية إلى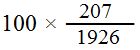 = 10,7٪ وكذلك ينتج التعابر المفرد بين e , sr الصنوف 5 , 6 , 7 , 8 ويكون مجموع أعداد التراكيب الجديدة لهذه الصنوف هو 183 أو 5,9٪ وكذلك ينتج التعابر المفرد بين e , cu الصنوف 3 , 4 , 5 , 6 ويكون مجموع أعداد التراكيب الجديدة لهذه الصنوف هو 384 أو 0,20 ٪ . إن حدوث التعابر المزدوج Double crossing over بين cu , e ترك نهاية النقاط في الترتيب الأبوي مما قلل المسافة بينهما . أن المسافة الصحيحة بين e ,cu على أساس التعابر المفرد هي 7,10 + 5,9 = 0,20 وحدة . تعتبر مسافة 10 وحدات أو أقل من ذلك حداً سليماً الذي لا يحدث فيه تعابر مزدوج في ذبابة الفاكهة . وعليه تبعد cu 7,10 وحدة مسافة عن sr وتبعد sr 5,9 وحدة مسافة عن e , وتبعد cu 0,20 وحدة مسافة عن e على الكروموسوم .
= 10,7٪ وكذلك ينتج التعابر المفرد بين e , sr الصنوف 5 , 6 , 7 , 8 ويكون مجموع أعداد التراكيب الجديدة لهذه الصنوف هو 183 أو 5,9٪ وكذلك ينتج التعابر المفرد بين e , cu الصنوف 3 , 4 , 5 , 6 ويكون مجموع أعداد التراكيب الجديدة لهذه الصنوف هو 384 أو 0,20 ٪ . إن حدوث التعابر المزدوج Double crossing over بين cu , e ترك نهاية النقاط في الترتيب الأبوي مما قلل المسافة بينهما . أن المسافة الصحيحة بين e ,cu على أساس التعابر المفرد هي 7,10 + 5,9 = 0,20 وحدة . تعتبر مسافة 10 وحدات أو أقل من ذلك حداً سليماً الذي لا يحدث فيه تعابر مزدوج في ذبابة الفاكهة . وعليه تبعد cu 7,10 وحدة مسافة عن sr وتبعد sr 5,9 وحدة مسافة عن e , وتبعد cu 0,20 وحدة مسافة عن e على الكروموسوم .
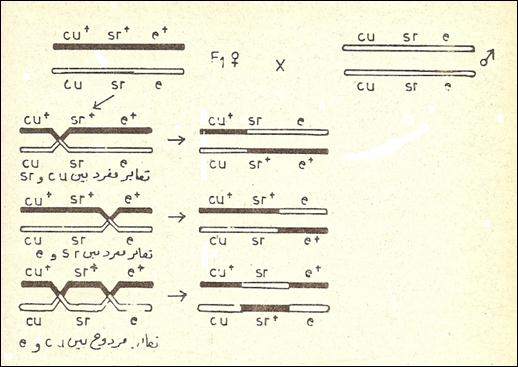
شكل (1) تضريب الثلاثة نقاط في الدروسوفلا وتعابرين مفردين ومعابر مزدوج .
ويمكن الاستفادة من هذه النتائج لإعداد جزء من الخريطة الكروموسوم الثالث الوراثية لذبابة الفاكهة . وقبل تحليل هذه النتائج لابد لنا أن نتذكر معلومات الهندسة المستوية حول ثلاثة نقاطc, b, a . فإذا كانت المسافة بين c , a مساوية إلى المسافة بين b , a مضافا أليها المسافة بين c , b يستنتج أن النقاط الثلاثة تقع على خط مستقيم أو تترتب ترتيباً خطياً . وعند تطبيق هذه المعلومات الهندسية على وحدات المسافة بين الجينات الثلاثة (e , sr , cu) نستنتج بأنها تترتب ترتيباً خطياً على الكروموسوم الثالث .
التداخل والتوافق Interference and Coincidence
لو كان توزيع التعابر في أي قسم من كروموسوم معين توزيعاً عشوائياً تاماً لأعطى احتمال حدوث تعابر بين مفردين بنفس الوقت في منطقتين متقاربتين حسب قانون الاحتمال بمقدار يساوي إلى ضرب احتمال حدوث التعابر المفرد في كل منطقة بصورة منفصلة . وأظهر مولر Muller عام 1916م بأن التعابر المزدوج الفعلي يكون أقل تردداً من المتوقع على أساس التوزيع العشوائي . وقدم مولر صيغة رياضية للتعبير عن هذه الظاهرة في ذبابة الفاكهة والذرة الصفراء . وتكون هذه الصيغة مساوية إلى النسبة بين تردد التعابر المزدوج المشاهد وتردد التعابر المتوقع أو تساوي إلى:

وسُمي هذا القياس الرياضي للتداخل بمعامل التوافق Coefficient of Coincidence وفي حالة التداخل التام لا يحدث أي تعابر مزدوج ويكون معامل التوافق مساوياً إلى الصفر . بينما في حالة غياب التداخل , يكون معامل التوافق مساوياً إلى الواحد . ويمكن استعمال معلومات تضريب الثلاثة نقاط في المثال المذكور أعلاه لحساب معامل التوافق الذي 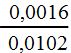 أي نتج 16٪ من التعابر المزدوج المتوقع والذي يشير إلى التداخل الجزئي بقيمة 84٪ . ويتوقع تداخل أقل في حالة كون الجينات متباعدة أكثر .
أي نتج 16٪ من التعابر المزدوج المتوقع والذي يشير إلى التداخل الجزئي بقيمة 84٪ . ويتوقع تداخل أقل في حالة كون الجينات متباعدة أكثر .
ويوجد حالة مختلفة في فطر الرشاسية Aspirgillus وعاثية البكتريا T4 ويظهر أن حدوث تعابر في منطقة ما يزيد احتمال حدوث تعابر آخر في المنطقة المجاورة , وعليه يكون معامل التوافق أكثر من واحد . وأطلق على هذه الظاهرة بالتداخل السلبي Negative interference .
خريطة الكروموسوم Chromosome Map
توفر عادة تضريبات الثلاثة نقاط معلومات لأعداد الخريطة الوراثية . وتمثل هذه الخرائط تخطيطاً للمواقع النسبية وللمسافات إلى الجينات في كل زمرة ارتباط . وعندما تراعى المسافات القصيرة (أقل من 10 وحدات) , يمكن استعمال إضافة خطية بسيطة عند التنقل من محل جين إلى محل جين آخر مجاور . وأعدت معظم المعلومات الضرورية لخريطة الكروموسوم الوراثية في العشرينيات من القرن الحالي من قبل علماء وراثة ذبابة الفاكهة وبالأخص من قبل موركان Morgan وبريجز Bridges وستورتيفانت Sturtevant . ومدرج في بعض الأماكن المعروفة جيداً للجينات ومواقعهم النسبية موزعين على زمر (مجاميع) الارتباط الأربعة المناظرة للأزواج الأربعة من الكروموسومات في ذبابة الفاكهة من نوع D. melanogaster من الواضح إن الزمر I , II , III تكون كبيرة وتحوي جينات كثيرة , أما الزمرة IV فإنها صغيرة جداً وتحوي جينات قليلة . وفي ذبابة الفاكهة أمكن إعداد الخرائط الخلوية للكروموسومات باستعمال معلومات من دراسات التي تربط بين التبدلات في النمط الظاهري بالتبدلات على الكروموسومات المتعددة الخيوط Polytene العملاقة .
وبالإضافة إلى الخرائط الوراثية لذبابة الفاكهة يوجد خرائط وراثية جيدة لكثير من النباتات كالذرة الصفراء والطماطا والشعير والحنطة والرز والذرة البيضاء والبزاليا . وبالمقارنة نجد إن التقدم البطيء في إعداد الخرائط الوراثية في اللبائن . وأعدت الخرائط الوراثية لتسعة عشر زوج من الكروموسومات للفأر المنزلي Mus musculus الحاوي على عشرين زوجاً من الكروموسومات بينما يكون إعداد الخرائط الكروموسومية للإنسان متعباً وغير منجز لكثير من الكروموسومات ولكن التقنيات الخلوية والإحصائية الجديدة أدت إلى تقدم كبير في هذا المجال .
العوامل التي تؤثر على ترددات التعابر Factors That Affect Crossover Frequencies
إن نسبة التعابر لا تكون ثابتة دائماً بين مكانين خاصين وقد تتغير بظروف التجارب المختلفة . يعتبر الجنس أحد العوامل التي تؤثر على كمية التعابر إلى درجة كبيرة أو قليلة . ومثال متطرف لهذا العامل هو ذكور ذبابة الفاكهة التي يكبت فيها التعابر تماماً . وعلى العموم يكون تردد التعابر في الجنس المتباين الأمشاج أوطأ مما عليه في الجنس المتماثل الأمشاج . وبالإضافة إلى الجنس . فيما يلي العوامل المهمة التي تؤثر على تردد التعابر في ذبابة الفاكهة :



|
|
|
|
"عادة ليلية" قد تكون المفتاح للوقاية من الخرف
|
|
|
|
|
|
|
ممتص الصدمات: طريقة عمله وأهميته وأبرز علامات تلفه
|
|
|
|
|
|
|
المجمع العلمي للقرآن الكريم يقيم جلسة حوارية لطلبة جامعة الكوفة
|
|
|
















