آخر المواضيع المضافة

 النبات
الحيوان
الأحياء المجهرية
علم الأمراض
التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحياتية النانوية
علم الأجنة
الأحياء الجزيئي
علم وظائف الأعضاء
الغدد
المضادات الحيوية
النبات
الحيوان
الأحياء المجهرية
علم الأمراض
التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحياتية النانوية
علم الأجنة
الأحياء الجزيئي
علم وظائف الأعضاء
الغدد
المضادات الحيوية| Morphology and Anatomy of Other Leaf Types |


|
|
Read More
Date: 13-11-2016
Date: 13-11-2016
Date: 14-11-2016
|
Morphology and Anatomy of Other Leaf Types
SUCCULENT LEAVES
Numerous adaptations permit plants to survive in desert habitats, one of the most common being the production of succulent leaves (Fig. 1). This is characteristic of species in the families Crassulaceae (contains Kalanchoe and Sedum), Portulacaceae (contains Portulaca and Lewisia), and Aizoaceae (ice plant), among others. The leaves are thick and fleshy, a shape that reduces the surface-to-volume ratio and favors water conservation. Some leaves are cylindrical or even spherical, the optimal surface-to-volume shape. The reduction in surface area, which is advantageous for water retention, has the automatic consequence of reducing the capacity for carbon dioxide uptake.

FIGURE 1 (a) Leaves of Senecio rotundifolia are spherical, giving them an optimal surface-to-volume ratio for co^T6ervi^:g water, (b) The leaves of the succulent Dinteranthus are hemispherical and attached to the opposite leaf, greatly reducing the exposed leaf surface. Interior tissues are mostly water- stage parenchyma. The low levels of chlorophyll result in low levels of photosynthesis, but lack of water is a more significant danger for these plants. Each plant consists basically of two leaves and a microscopic stem; the root system (not visible) is substantial. Dinteranthus and many related spades have only two leaves at a time; as two new leaves expand, the existing two collapse and wither away.
Inside the leaf, the mesophyll contains very few air spaces, reducing the internal evaporative surface area and, in turn, water loss through stomata. Lack of air spaces also makes the leaves more transparent, just as pure water is more transparent than soap bubbles or foam, allowing light to penetrate farther into the leaf. Photosynthesis can occur much more deeply than it would in the foliage leaves described earlier. In some members of the genera Lithops (stone plants) and Frithia, leaves are so translucent that they act as optical fibers; the leaves are located almost completely underground, where it is cool and relatively damp (Fig. 2). The exposed leaf tips allow sufficient light to enter and be conducted to the subterranean chlorenchyma. Although the plants live in a harsh desert in Madagascar, photosynthesis actually occurs in a rather mild microclimate.

FIGURE 2: Lithops, stone plant, is closely related to Dinteranthus; both are in the family Aizoaceae. Lithops also has only two leaves; but they are located almost entirely underground. The flat, translucent tips project above the soil and conduct light to the subterranean chlorenchyma.
SCLEROPHYLLOUS FOLIAGE LEAVES
Foliage leaves must produce more sugars by photosynthesis than are used in their own construction and metabolism, or the plant would lose energy every time it produced a leaf. This requirement limits the amount of sclerenchyma in foliage leaves, so most leaves tend to be soft, flexible, and edible. In some species (barberry, holly, Yucca), leaves have evolved which are perennial, existing on a plant for two or more years (3). With this extended lifetime and increased productivity, sclerenchymatous leaves are feasible, and their hardness makes them more resistant to animals, fungi, freezing temperatures, and ultraviolet light. Such plants are said to be sclerophyllous, and the leaves are sclerophylls. The scler- enchyma is often present as a layer just below the epidermis and in the bundle sheaths, although the epidermis itself can be composed of thick-walled cells (Fig. 4). The cuticle is usually very thick, and waxes are abundant on leaves of many sclerophyllous species.

FIGURE 3: The leaves of barberry (Berberis) are tough and hard; it is difficult for insects to bite into them or lay eggs in them. However, the plant must invest considerable glucose to make the sclerenchyma cells, so the leaves must photosynthesize longer before they reach the break-even point
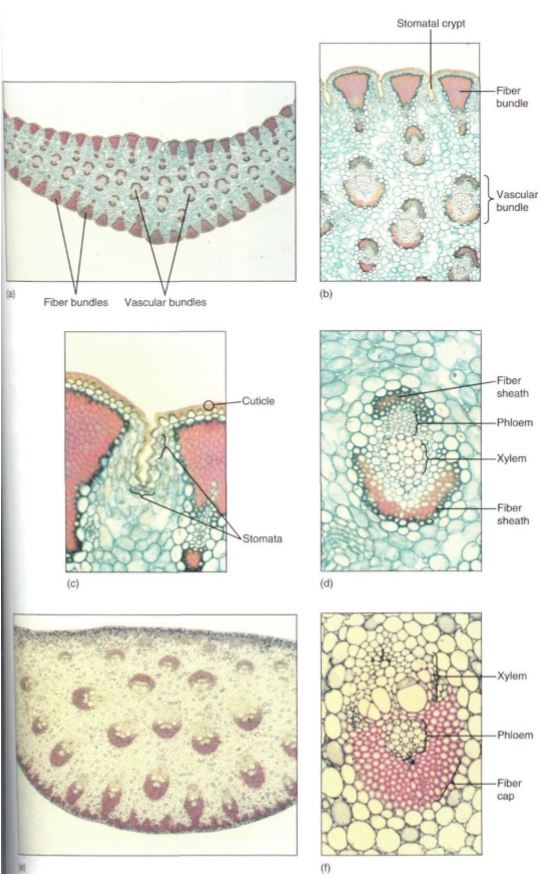
FIGURE 4: (a) Yucca leaves are excellent examples of sclerophylls; they are tough and fiberous, and the cuticle is thick. These features reduce the amount of light reaching the chlorenchyma, but plant growth is probably limited by scarcity of water rather than reduced photosynthesis. These leaves may function for 5 to 10 years or more (X 15). (b) Bundles of fibers occur just interior to the epidermis, and the thick mesophyll surrounds many vascular bundles (X 50). (c) Stomata occur in long, shallow grooves that run between the fiber bundles (X 150). (d) Even the vascular bundles of these sclerophylls have fiber sheaths (X 150). (e) The sclerophylls of Dracaena are very thick and have leaf veins throughout the mesophyll, unlike the single row in ordinary leaves (X 15). (f) The leaf veins of Dracaena have extremely thick fiber sheaths (X 150).
LEAVES OF CONIFERS
In almost all species of conifers, the leaves are sclerophylls; they have an extremely thick cuticle and the cells of their epidermis and hypodermis have thick walls. Most confer leaves contain abundant chemicals that make them unpalatable. Conifer leaves are always simple and have only a few forms. Needles, either short or long, occur in all pines, firs, and spruces (Fig. 5). Needles of longleaf pine can be 40 cm long, although most ether species of pine have needles about 10 cm long. Small, flat, scale-like leaves form a shieldlike covering on stems of junipers, cypresses (Cupressus), arborvitae (Thuja), and others. In Agathis, Araucaria, and Podocarpus, all genera of the Southern Hemisphere, leaves are rather large, broad scales held away from the stem. Lengths up to 12.5 cm and widths of 3.5 cm have been measured in leaves of Podocarpus wallichianus.

FIGURE 5: (a) Many conifers, such as this pine, have needle-shaped leaves. (b) Incense cedar (Libocedrus) and many other conifers have scale-shaped leaves.
Conifer leaves are mostly perennial, remaining on the stem for many years; cons- quently, the plants are evergreens. Needles of bristlecone pine live for at least 5 years; their vascular bundles can produce new phloem each year, but no new xylem (Fig. 6). The small scale leaves of juniper also persist and are photosynthetically active for many years, and leaves of Agathis and Araucaria remain even on very old trunks. Some conifers have annual leaves that are shed each autumn; larch (Larix), bald cypress (Taxodium), and dawn redwood (Metasequoia) are all deciduous.

FIGURE 6: (a) The needle leaves of pine are round in transverse section (X 15). (b) The needles last for many years, producing more phloem each year (X 150). (c) Pine needles are sclerophylls: they have thickwalled epidermal cells with a thick cuticle. They also contain resin canals (X 150). (d and e) Leaves of yew (Taxus) are flat, but they also have sclerophyllous features and produce phloem year after year (d, X50;e, X 150).
BUD SCALES
One of the most common modifications of leaves is their evolutionary conversion into bud scales (7). In perennial plants, dormant shoot apical meristems must be protected from low temperatures and the drying action of wind during winter. Bud scales provide this protection by forming a tight layer around the stem tip. Because their role is primarily protection, not photosynthesis, they have a different structure from foliage leaves. Bud scales are small and rarely compound, so mechanical wind damage is not a risk for bud scales. Their petiole is either short or absent because they must remain close to the stem and be folded over it. To be protective, they must be tougher and more waxy than regular leaves; bud scales frequently produce a thin layer of corky bark, at least on exposed portions, which provides greater protection than the simple epidermis of foliage leaves.

FIGURE 7 :(a) Bud scales of Rhododendron fit together tightly during winter, protecting the enclosed apical meristem and young leaves and flowers. (b) The bud scales of Magnolia have a dense covering of hair; this improves the insulation.
SPINES
Many spines are modified leaves, especially in the cacti that grow in arid regions. The succulent, moist cactus body would be an excellent source of water for animals were it not for the protective spines (Fig. 8). As with bud scales, spines have a distinct structure related to their function. The soft, flexible blade of a photosynthetic leaf is generally useless as a protective device against herbivores, but spines have no blade and are needle-shaped; mutations that inhibit lamina formation have been selectively advantageous. No mesophyll parenchyma or vascular tissue is present; the mesophyll instead consists of closely packed fibers. Once fibers mature, they deposit lignin in their walls, which makes them hard and resistant to decay. The cells then die and dry out, hardening even further. Because cacti carry out photosynthesis in their stem cortex, loss of the leaf lamina was selectively advantageous. But for plants that have no alternative photosynthetic tissues, mutations that cause loss of lamina are extremely disadvantageous.
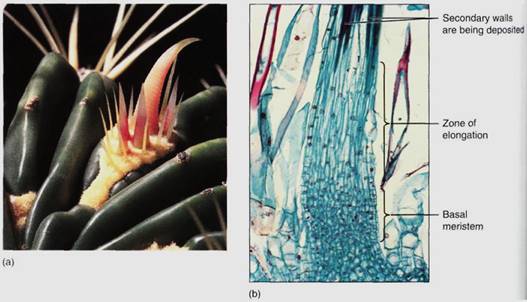
FIGURE 8: (a) The spines of this barrel cactus (Fervcactus) are modified leaves. The cluster of spines is actually an entire axillary bud. (b) Cactus spines have a basal meristem; as new cells are formed, older cells are pushed upward. As they move out of the meristem, they fill their central vacuole with water, elongate, then deposit a thick secondary wall and differentiate into fiber cells (X 50).
TENDRILS
The tendrils of many plants (peas, cucumbers, and squash) are another form of modified leaf (Fig. 9). Unlike photosynthetic leaves, tendrils grow indefinitely and contain cells capable of sensing contact with another object. When the tendril touches something, the side facing the object stops growing but the other side continues to elongate, causing the tendril to coil around the object and use it for support. A lamina would be detrimental, and none forms. Whereas many foliage leaves are capable of sensing the direction of sunlight and reorienting the lamina for maximum photosynthesis, tendrils respond by sensing solid objects and growing around them.

FIGURE 9: This tendril is a highly modified leaf: It has no leaf blade, and whereas foliage leaves stop growing after they reach a specific size, this tendril continues to grow.
LEAVES WITH KRANZ ANATOMY
A distinct type of leaf anatomy occurs in plants that have a special metabolism called C4 . These plants lack a palisade parenchyma and spongy mesophyll but have prominent bundle sheaths composed of large chlorophyllous cells. Surrounding each sheath is a ring of mesophyll cells that appear to radiate from the vascular bundle. These plants possess a mechanism of carbon dioxide transport that requires this special Kranz anatomy and adapts C4 plants to arid environments.
INSECT TRAPS
The ability to trap and digest insects has evolved in several families. Insectivory has evolved in plants that grow in habitats poor in nitrates and ammonia; by digesting insects, plants obtain the nitrogen they need for their amino acids and nucleotides. Trap leaves can be classified as either active traps that move during capture or passive traps incapable of movement. The best-known passive traps are the pitcher-leaves of Nepenthes, Darlingtonia, and Sarracenia (Fig. 10a). Although the leaf appears highly modified, it is actually quite similar to many foliage leaves. It is thin, parenchymatous, and capable of photosynthesis. It has numerous stomata and vascular bundles as well as a mesophyll containing aerenchyma and chlorenchyma. The most significant differences are that the lamina is tubular rather than flat and that it secretes a watery digestive fluid. The epidermis in the digestive region must be absorptive rather than impermeable. Also, the throat of the pitcher contains numerous trichomes that point toward the liquid; it is easier for insects to walk in the direction of the trichomes, and they are thus led to their death.
The leaves of sundew (Drosera; Fig.10b) are active traps; they too have many features in common with foliage leaves, but their upper surface is covered with glandular trichomes that secrete a sticky digestive liquid. Once an insect is caught on a single trichome, adjacent trichomes are stimulated to bend toward the victim, placing their digestive drops on it as well. The entire leaf blade curls around the insect so that many trichomes come into contact with it. The modifications in these leaves have been largely metabolic rather than structural: Trichomes and lamina must be able to sense and respond to the presence of an insect, carry on secretion and absorption, and unfold after digestion is complete.
Venus' flytraps (Dionaea muscipula) have leaves that are held flat, like most foliage leaves, but in Dionaea this position is maintained only because motor cells along the upper side of the midrib are extremely turgid and swollen (Fig. 10c). When an insect walks across the trap, it brushes against trigger hairs. If two of these are stimulated, they cause the midrib motor cells to lose water quickly, and the trap rapidly closes as the two halves of the lamina move upward. On the margins are long interdigitating teeth that trap the insect; short glands begin to secrete digestive liquid. Once digestion and absorption are complete, the midrib motor cells fill with water, swell, and force the trap open, ready for a new victim.

FIGURE 10 :(a) Leaves of Nepenthes are the most elaborate of the pitcher leaves; they have an ordinary petiole and a broad lamina, but the leaf tip is long, narrow, and pendant. Its extreme tip turns upright and develops into the hollow pitcher that contains a digestive mixture and an epidermis that can absorb nitrogen. The ultimate portion of the leaf tip is a broad flat roof that prevents rain from fallirg into the pitcher and diluting the digestive juices. (b) Sundew leaves have many shapes; toe of Drosera capensis are long and narrow. Just after an insect is caught, the leaf curls and paos numerous tentacles (trichomes) on it. The drop at the end of each tentacle is both sticky and digestive. Once digestion is complete, the leaf uncurls and is ready for the next meal. (c) Each half of the blade of a Venus' flytrap leaf has three hairs; if an insect touches any two, it stimulates the trap to close. (c, Ray Coleman/Photo Researchers)



|
|
|
|
"إنقاص الوزن".. مشروب تقليدي قد يتفوق على حقن "أوزيمبيك"
|
|
|
|
|
|
|
الصين تحقق اختراقا بطائرة مسيرة مزودة بالذكاء الاصطناعي
|
|
|
|
|
|
|
قسم شؤون المعارف ووفد من جامعة البصرة يبحثان سبل تعزيز التعاون المشترك
|
|
|