آخر المواضيع المضافة

 النبات
الحيوان
الأحياء المجهرية
علم الأمراض
التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحياتية النانوية
علم الأجنة
الأحياء الجزيئي
علم وظائف الأعضاء
الغدد
المضادات الحيوية
النبات
الحيوان
الأحياء المجهرية
علم الأمراض
التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحياتية النانوية
علم الأجنة
الأحياء الجزيئي
علم وظائف الأعضاء
الغدد
المضادات الحيوية| The MHC Locus Comprises a Cohort of Genes Involved in Immune Recognition |


|
|
Read More
Date: 14-12-2015
Date: 25-3-2021
Date: 9-6-2021
|
The MHC Locus Comprises a Cohort of Genes Involved in Immune Recognition
KEY CONCEPTS
- The MHC locus encodes class I, class II, and class III molecules. Class I proteins are the transplantation antigens distinguishing “self” from “nonself.” Class II proteins are involved in interactions of T cells with antigen-presenting cells (APCs). Class III molecules are diverse and include cytokines and components of the complement cascade.
- MHC class I molecules are heterodimers consisting of a variant α chain and the invariant β -microglobulin.
- MHC class II molecules are heterodimers consisting of an α chain and a β chain.
MHC molecules have evolved to maximize the efficacy and flexibility of their function: to bind peptides derived from microbial pathogens and present them to T cells. In response to a strong evolutionary pressure to eliminate a large variety of microorganisms, the MHC genes encoding these proteins have evolved into polygenic (several sets of genes in all individuals) and polymorphic (multiple variants of gene within the population at large) cohorts of genes. In humans, the MHC is also called human leukocyte antigen (HLA). MHC proteins are dimers inserted in the plasma membrane, with a major part of the protein protruding on the extracellular side. Of the three human MHC classes, class I and class II are the most important in immunobiology and the clinical setting. The structures of MHC class I and class II molecules are related, although they are made up of different components (FIGURE 1).

FIGURE 1.Class I and class II MHC molecules have a related structure. Class I antigens consist of a single polypeptide (α) with three external domains (α1, α2, and α3) that interacts with β2 -microglobulin (β2M). Class II antigens consist of two polypeptides (α and β), each with two domains (α1 and α2 and β1 and β2) with a similar overall structure.
MHC class I molecules consist of a heterodimer of the class I chain (α) itself and the β2-microglobulin (β2M protein). The class I chain is a 45-kD transmembrane component that has three external domains (each approximately 90 amino acids long), one of which interacts with β2-microglobulin, a transmembrane domain (approximately 40 residues), and a short cytoplasmic domain (approximately 30 residues). MHC class II molecules consist of two chains, α and β, whose combination generates an overall structure in which there are two extracellular domains. Humans have three classified (or major) class Iα-chain genes: HLA-A, HLA-B, and HLA-C. The β2-microglobulin is a secreted protein of 12 kD. It is needed for the class I chain to be transported to the cell surface.
Mice lacking the β2-microglobulin gene express no MHC class I antigens on the cell surface. Humans have three major pairs of class IIα- and β-chain genes: HLA-DR, HLA-DP, and HLA-DQ.
The MHC locus occupies a small region of a single chromosome in mice (histocompatibility 2 or H2 locus on chromosome 17) and in humans (human leukocyte antigen or HLA locus on chromosome 6). These regions contain multiple genes. Also located in these regions are genes encoding proteins found on lymphocytes and macrophages that have a related structure and are important in the function of cells of the immune system.
The genes of the MHC locus are grouped into three clusters according to the structures and immunological properties of the respective products. The MHC region was originally defined by genetics in the mouse, where the classical H2 region occupies 0.3 map units. Together with the adjacent region, where mutations affecting immune function are also found, this corresponds to an approximately 2,000-kb region. The MHC region is generally conserved in mammals, as well as in some birds and fish. The genomic regions where the class I and class II genes are located mark the original boundaries of the locus, from telomere to centromere (FIGURE 2: right to left). The genes in the class III region, which separate class I from class II genes, encode many proteins with a variety of functions. Defining the ends of the locus varies with the species; the area beyond the class I genes on the telomeric side is called the extended class I region. Likewise, the region beyond the class II gene cluster on the centromeric side is referred to as extended class II region. The major difference between mice and humans is that the extended class II region contains some class I (H2-K) genes in mice.

FIGURE 2.The MHC region extends for more than 2 Mb. MHC proteins of classes I and II are encoded by two separate regions. The class III region is defined as the segment between them. The extended regions describe segments that are syntenic on either end of the cluster. The major difference between mouse and human is the presence of H2 class I genes in the extended region on the left. The murine locus is located on chromosome 17, and the human locus is located on chromosome 6.
The organization of class I genes is based on the structure of their products (Figure 3). The first exon encodes a signal sequence, cleaved from the protein during membrane passage. The next three exons encode each of the external domains. The fifth exon encodes the transmembrane domain. The last three rather small exons together encode the cytoplasmic domain. The only difference in the genes for human transplantation antigens is that their cytoplasmic domain is coded by only two exons. The exon encoding the third external domain of the class I genes is highly conserved relative to the other exons. The conserved domain probably represents the region that interacts with β2-microglobulin, which explains the need for constancy of structure. This domain also exhibits homologies with the constant region domains of Igs.
Most of the sequence variation between class I alleles occurs in the first and second external domains, sometimes taking the form of a cluster of base substitutions in a small region.
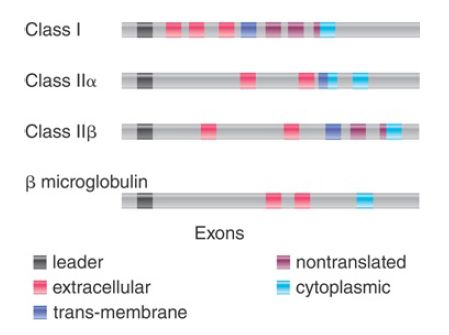
FIGURE 3.Each class of MHC genes has a characteristic organization, in which exons represent individual protein domains.
The gene for β2-microglobulin is located on a separate chromosome. It has four exons, the first encoding a signal sequence, the second encoding the bulk of the protein (from amino acids 3 to 95), the third encoding the last four amino acids and some of the nontranslated UTR, and the last encoding the rest of the UTR. The length of β2-microglobulin is similar to that of an Ig V gene; there are certain similarities in amino acid constitution, and there are some (limited) homologies of nucleotide sequence between β2-microglobulin and Ig constant domains or type I gene third external domains.
MHC class I genes encode transplantation antigens. They are present on every mammalian cell. As their name suggests, these proteins are responsible for the rejection of foreign tissue, which is recognized as such by virtue of its particular array of transplantation antigens. In the immune system, their presence on target cells is required for cell-mediated responses. The types of class I proteins are defined serologically by their antigenic properties. The murine class I genes encode the H2-K and H2-D/L proteins. Each mouse strain has one of several possible alleles for each of these proteins. The human class I genes encode the classical transplantation antigens: HLA-A, HLA-B, and HLA-C.
Some HLA class I–like genes lie outside the MHC locus. Notable among these genes are those of the small CD1 family. CD1 genes encode proteins expressed on DCs and monocytes. CD1 proteins can bind glycolipids and present them to T cells, which are neither CD4 nor CD8.
MHC class II genes encode the MHC class II proteins. These are expressed on the surfaces of both B and activated T lymphocytes, as well as on macrophages and dendritic cells. MHC class II molecules are critically involved in antigen presentation and communications between cells that are necessary to induce a specific immune response. In particular, they are required for T cell function. The murine class II genes were originally identified as immune response (Ir) genes; that is, genes whose expression made it possible for an immune response to a given antigen to be triggered (hence, the I-A and I-E terminology). The human class II region (also called HLA-D) is arranged into HLA-DR, HLA-DP, and HLA-DQ subregions. This region also includes several genes that are related to the initiation of antigen-specific response, namely, antigen presentation. These genes include those encoding TAP and LMP, as well as those encoding the DM and DO molecules, which regulate peptide loading onto classical class II molecules.
Expression of nonclassical MHC class II is induced by IFN-γ through CIITA, the MHC class II transcriptional activator. MHC class III genes occupy the “transitional” region between class I and class II regions. The class III region includes genes encoding complement components, including C2, C4, and factor B. The role of complement factors is to interact with antibody–antigen complexes and mediate activation of the complement cascade, eventually lysing cells, bacteria, or viruses. Other genes lying in this transitional region include those encoding tumor necrosis factor-α (TNF-α) and lymphotoxin-α (LTA) and lymphotoxin-β (LTB).
The MHC regions of mammals have several hundred genes, but it is possible for MHC functions to be provided by far fewer genes, as in the case of chickens, where the MHC region is 92 kb andcomprises only 19 genes. In compar ison to other gene families, the
exact numbers of genes devoted to each function differs. The MHC locus shows extensive variation between individuals, and a number of genes may be different in different individuals. As a general rule, however, a mouse genome has fewer active H2 genes than a human genome. The class II genes are unique to mammals (except for one subgroup); birds and fish have different genes in their place. Humans have approximately 8 functional class I genes; mice have approximately 30. The class I region also includes many other genes. The class III regions are very similar in humans and mice.
MHC class I and class II genes are highly polymorphic, with the exception of human HLA-DRα and the mouse homologue H2-Eα, and likely arose as a result of extensive gene duplications. Further divergence arose through mutations and gene conversion.



|
|
|
|
التوتر والسرطان.. علماء يحذرون من "صلة خطيرة"
|
|
|
|
|
|
|
مرآة السيارة: مدى دقة عكسها للصورة الصحيحة
|
|
|
|
|
|
|
نحو شراكة وطنية متكاملة.. الأمين العام للعتبة الحسينية يبحث مع وكيل وزارة الخارجية آفاق التعاون المؤسسي
|
|
|