
آخر المواضيع المضافة

 النبات
الحيوان
الأحياء المجهرية
علم الأمراض
التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحياتية النانوية
علم الأجنة
الأحياء الجزيئي
علم وظائف الأعضاء
الغدد
المضادات الحيوية
النبات
الحيوان
الأحياء المجهرية
علم الأمراض
التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحياتية النانوية
علم الأجنة
الأحياء الجزيئي
علم وظائف الأعضاء
الغدد
المضادات الحيوية| Viral DNA Is Generated by Reverse Transcription |


|
|
Read More
Date: 7-4-2021
Date: 28-4-2016
Date:
|
Viral DNA Is Generated by Reverse Transcription
KEY CONCEPTS
- A short sequence (R) is repeated at each end of the viral RNA, so the 5′ and 3′ ends are R-U5 and U3-R, respectively.
- Reverse transcriptase starts synthesis when a tRNA primer binds to a site 100 to 200 bases from the 5′ end.
- When the enzyme reaches the end, the 5′ terminal bases of RNA are degraded, exposing the 3′ end of the DNA product.
- The exposed 3′ end of the DNA product base pairs with the 3′ terminus of another RNA genome.
- Synthesis continues, generating a product in which the 5′ and 3′ regions are repeated, giving each end the structure U3-R-U5.
- Similar strand-switching events occur when reverse transcriptase uses the DNA product to generate a complementary strand.
- Strand switching is an example of the copy choice mechanism of recombination.
Retroviruses are called plus-strand viruses, because the viral RNA itself codes for the protein products. As its name implies, reverse transcriptase is responsible for converting the genome (plus-strand RNA) into a complementary DNA strand, which is called the minus-strand DNA. Reverse transcriptase also catalyzes subsequent stages in the production of duplex DNA. It has a DNA polymerase activity, which enables it to synthesize a duplex DNA from the single-stranded reverse transcript of the RNA.
The second DNA strand in this duplex is called the plus-strand DNA. As a necessary adjunct to this activity, the enzyme has an RNase H activity, which can degrade the RNA part of the RNA–DNA hybrid. All retroviral reverse transcriptases share considerable similarities of amino acid sequence, and homologous sequences can be recognized in all other retroelements.
The structures of the DNA forms of the virus are compared with the RNA in FIGURE 1. The viral RNA has direct repeats at its ends. These R segments vary in different strains of virus, ranging from 10 to 80 nucleotides. The sequence at the 5′ end of the virus is R-U5, and the sequence at the 3′ end is U3-R. The R segments are used during the conversion from the RNA to the DNA form to generate the more extensive direct repeats that are found in linear DNA, as shown in FIGURE 2and FIGURE 3. The shortening of 2 bp at each end in the integrated form is a consequence of the mechanism of integration .
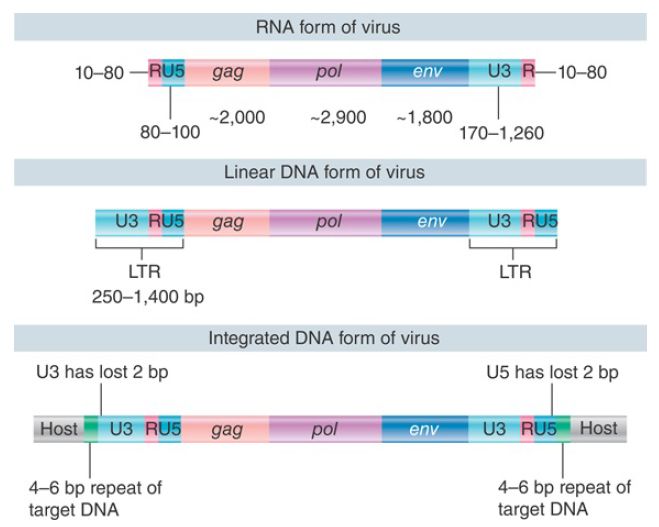
FIGURE 1. Retroviral RNA ends in direct repeats (R), the free linear DNA ends in LTRs, and the provirus ends in LTRs that are shortened by two bases each.
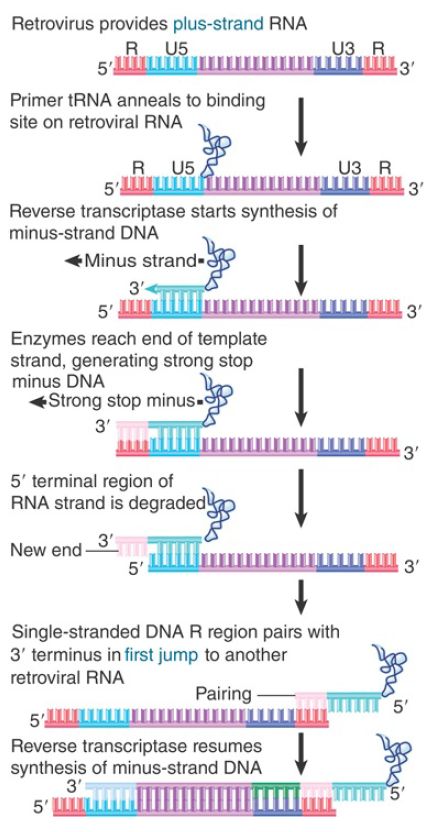
FIGURE 2. Minus-strand DNA is generated by switching templates during reverse transcription.
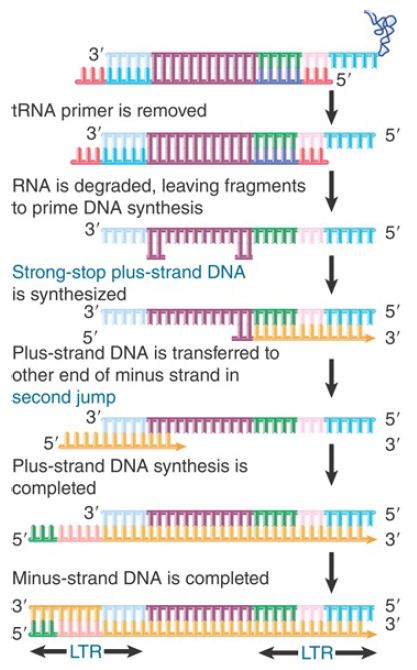
FIGURE 3. Synthesis of plus-strand DNA requires a second jump.
Like other DNA polymerases, reverse transcriptase requires a primer. For retroviruses, the native primer is tRNA. An uncharged host tRNA is present in the virion. A sequence of 18 bases at the 3′ end of the tRNA is base paired to a site 100 to 200 bases from the 5′ end of one of the viral RNA molecules. The tRNA may also be base paired to another site near the 5′ end of the other viral RNA, thus assisting in dimer formation between the viral RNAs.
Here is a dilemma: Reverse transcriptase starts to synthesize DNA at a site only 100 to 200 bases downstream from the 5′ end. How can DNA be generated to represent the intact RNA genome?
Synthesis in vitro proceeds to the end, generating a short DNA sequence called strong-stop minus DNA. This molecule is not found in vivo because synthesis continues by the reaction illustrated in Figure 3. Reverse transcriptase switches templates, carrying the nascent DNA with it to the new template. This is the first of two jumps between templates.
In this reaction, the R region at the 5′ terminus of the RNA template is degraded by the RNase H activity of reverse transcriptase. Its removal allows the R region at a 3′ end to base pair with the newly synthesized DNA. Reverse transcription then continues through the U3 region into the body of the RNA.
The source of the R region that pairs with the strong-stop minus DNA can be either the 3′ end of the same RNA molecule (intramolecular pairing) or the 3′ end of a different RNA molecule (intermolecular pairing). The switch to a different RNA template is used in the figure because evidence suggests that the sequence of the tRNA primer is not inherited in a retroposon life cycle. (If intramolecular pairing occurred, we would expect the sequence to be inherited, because it would provide the only source for the primer binding sequence in the next cycle. Intermolecular pairing allows another retroviral RNA to provide this sequence.)
The result of the switch and extension is to add a U3 segment to the 5′ end. The stretch of sequence U3-R-U5 is called the long terminal repeat (LTR) because a similar series of events adds a U5 segment to the 3′ end, giving it the same structure of U3-R-U5. Its length varies from 250 to 1,400 bp (see Figure 1). We now need to generate the plus strand of DNA and to generate the LTR at the other end. The reaction is shown in Figure 3.
Reverse transcriptase primes synthesis of plus-strand DNA from afragment of RNA that is left after degrading the original RNA molecule. A strong-stop plus-strand DNA is generated when the enzyme reaches the end of the template. This DNA is then transferred to the other end of a minus strand, where it is probably released by a displacement reaction when a second round of DNA synthesis occurs from a primer fragment farther upstream (to its left in the figure). It uses the R region to pair with the 3′ end of a minus-strand DNA. This double-stranded DNA then requires completion of both strands to generate a duplex LTR at each end.
Each retroviral particle carries two RNA genomes. This makes it possible for recombination to occur during a viral life cycle. In principle this could occur during minus-strand synthesis and/or during plus-strand synthesis:
The intermolecular pairing shown in Figure 2 allows a recombination to occur between sequences of the two successive RNA templates when minus-strand DNA is synthesized. Retroviral recombination is mostly due to strand transfer at this stage, when the nascent DNA strand is transferred from one RNA template to another during reverse transcription.
Plus-strand DNA may be synthesized discontinuously, in a reaction that involves several internal initiations. Strand transfer during this reaction can also occur, but is less common.
The common feature of both events is that recombination results from a change in the template during the act of DNA synthesis. This is a general example of a mechanism for recombination called copy choice. For many years this was regarded as a possible mechanism for general recombination. It is unlikely to be employed by cellular systems, but it is a common basis for recombination during infection by RNA viruses, including those that replicate exclusively through RNA forms, such as poliovirus.
Strand switching occurs with a certain frequency during each cycle of reverse transcription; that is, in addition to the transfer reaction that is forced at the end of the template strand. The principle is illustrated in FIGURE 4, although not much is known about the precise mechanism. Reverse transcription in vivo occurs in a ribonucleoprotein complex, in which the RNA template strand is bound to virion components, including the major protein of the capsid. In the case of human immunodeficiency virus (HIV), addition of this protein (NCp7) to an in vitro system causes recombination to occur. The effect is probably indirect: NCp7 affects the structure of the RNA template, which, in turn, affects the likelihood that reverse transcriptase will switch from one template strand to another.
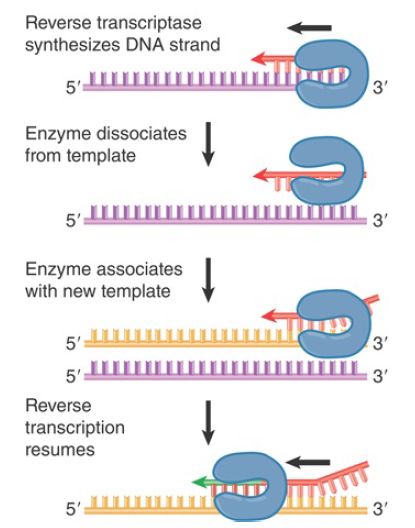
FIGURE 4. Copy choice recombination occurs when reverse transcriptase releases its template and resumes DNA synthesis using a new template. Transfer between template strands probably
occurs directly, but is shown here in separate steps to illustrate the process.



|
|
|
|
"عادة ليلية" قد تكون المفتاح للوقاية من الخرف
|
|
|
|
|
|
|
ممتص الصدمات: طريقة عمله وأهميته وأبرز علامات تلفه
|
|
|
|
|
|
|
المجمع العلمي للقرآن الكريم يقيم جلسة حوارية لطلبة جامعة الكوفة
|
|
|
















