آخر المواضيع المضافة

 النبات
الحيوان
الأحياء المجهرية
علم الأمراض
التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحياتية النانوية
علم الأجنة
الأحياء الجزيئي
علم وظائف الأعضاء
الغدد
المضادات الحيوية
النبات
الحيوان
الأحياء المجهرية
علم الأمراض
التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحياتية النانوية
علم الأجنة
الأحياء الجزيئي
علم وظائف الأعضاء
الغدد
المضادات الحيوية| Division Bryophyta: Mosses |


|
|
Read More
Date: 16-11-2016
Date: 15-11-2016
Date: 17-11-2016
|
Division Bryophyta: Mosses
THE GAMETOPHYTE GENERATION
Mosses are ubiquitous, occurring in all parts of the world and in almost every environment (Table ). They are perennial and thrive in many places within cities (Tortula on walls, Mnium on soil).
Morphology. The leafy stems, technically known as gametophores, of many moss plants grow close together, tightly appressed and forming dense mounds (Grimmia, Pohlia). In other species, particularly those of cool wet areas, the plants are more open and loose (Anamla, Climacium, Platygyrium;). Scouleria gametophores grow as ribbons up to 15 cm long, submerged in rapidly flowing water. Moss plants have stems, usually very short, and almost microscopic leaves (Fig. 1). Because the stems and leaves of mosses are parts of a gametophyte, not a sporophyte, they are not homologous with those of vascular plants. That is, the leaves of mosses did not evolve from the same structures as vascular plant stems and leaves did.
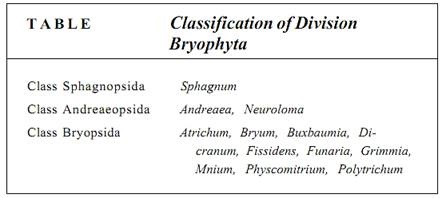

FIGURE 22.8:The individual shoots of a moss superficially resemble those of a flowering plant, having stems, leaves, nodes and internodes, and even buds. None of these structures is homologous to the equivalent organ in flowering plants, but the same set of names is used for both groups. Dark green, haploid tissue; light green, diploid tissue.
Moss gametophores grow from an apical meristem that contains a prominent apical cell (Fig. 2a). Derivative cells subdivide, producing the tissues of stem and leaves in rather precise arrangements. Leaves are aligned in three rows on the stem, at least while young; later expansion may obscure this pattern. Moss leaves are broad and most have a midrib (costa). The leaves of almost all mosses are only one cell thick except at the midrib and along the margin (Fig. 3a). In the family Polytrichaceae, common along roadsides in forested areas of the northern United States, genera such as Polytrichum and Atrichum have thin lamellae on the leaf upper surface, greatly increasing the photosynthetic 1k9le (Fig. 3b). Cuticle occurs only on the upper surface of most moss leaves, the underside being uncutinized and capable of absorbing water directly from rain, dew, and fog. Wafer is usually not transported into leaves from stems, because mosses are nonvascular plants. The lack of a cuticle on the lower or both sides of leaves means that mosses have little protection against desiccation; when the microhabitat dries, so do the leaves. No stomata occur on moss leaves; they would be useless because the leaves are unistratose.
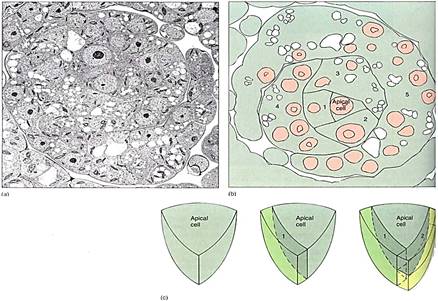
FIGURE 2: (a and b) Cross-section of the shoot apex of the moss Physcomitrium pyriforme: The apical cell is in the center, and its most recent daughter cell is to the left (#1). The next older cell (#2) has subdivided, as have successively older derivatives (#3, 4, 5) (X 2400). (c) The apical cell is shaped like an inverted pyramid. (Courtesy of B. Fulginiti, University of Texas).
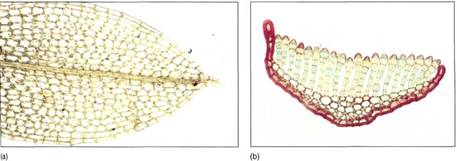
FIGURE 3: (a) Many moss leaves are only one cell thick, but the leaf "midrib" is thicker and is true parenchyma. (b) In the family Polytrichaceae, leaves bear long sheets of cells on their upper surface; this greatly increases the volume of photosynthetic tissue but does not decrease the high surface-to-volume ratio as it would if this were a solid tissue (X 200).
Moss stems are always slender and have little tissue differentiation (Fig. 4). The surface layer is at most only slightly different from the underlying layers and is not called epidermis. The stem tissues, all called cortex, may be uniform in all parts, or the outer cells nay be slightly narrower with walls that are somewhat thickened. Inner cells are larger, more parenchymatous, and chlorophyllous. In a few species of mosses, stems have hairs, but stomata do not occur.

FIGURE 4:Although most moss stems do not have vascular tissues, they do support the shoot, and most have a layer or two of thick-walled cells. Sugars and minerals must be transported from leaves to the foot apex, gametangia, and sporophytes, so living parenchyma cells are necessary (a, X 16; b, X 40).
Water Transport. In some mosses, primarily the family Polytrichaceae, the innermost cortex is composed of conducting cells called hydroids, elongated cells that lose their cytoplasm when mature (Fig. 5). Their end walls are partially digested and become more permeable to water and dissolved solutes, but they are not removed entirely. Each hydroid is aligned with those above and below it. If a moss is given a soluble form of lead (Pb-EDTA) and then later treated with hydrogen sulfide, a lead precipitate is found in the hydroids. This indicates that hydroids do conduct water and dissolved minerals.
Species that have hydroids typically also have leptoids, cells that resemble sieve cells (Fig. 5b). They are elongate, have relatively prominent interconnections with adjacent cells, and lack nuclei at maturity, although they do retain their cytoplasm. Adjacent parenchyma cells are unusually cytoplasmic and rich in enzymes, just as are companion cells. If radioactive bicarbonate is placed on the leaf, radioactive sucrose can be found later in the leptoids, moving as rapidly as 32 cm/hr. In the mosses that do not have leptoids, sugar is moved simply between parenchyma cells by slow transport.

FIGURE 5: (a) Hydroids lose their cytoplasm at maturity, just as do tracheids and vessel elements. Hydroids, however, do not have specialized secondary walls. (b) Leptoids are analogous to sieve cells, phloem cells that are simpler than sieve tube members. All pores are small, and the cells remain alive at maturity. (c) Face view of a leptoid end wall with plasmodesmata. (Courtesy of Dan Scheirer, Northeastern University).
In the majority of mosses, which lack hydroids and leptoids, water is conducted along the exterior of the plant by capillary action. Leaves and stems are so small that they form spaces narrow enough to act as capillary channels and transport water. In some species, such as Funaria, the leaves curl and become more closely appressed to the stem as they dry (Fig. 6); when rain or dew returns, the dried plant has even more capillary spaces than in the hydrated condition when the leaves are spread away from the stem.
At the base of the stem or along the length of horizontal stems are rhizoids. These small, multicellular trichome-like structures penetrate the surface of the substrate. Rhizoids only anchor the stem; they do not appear to be involved in absorbing either water or minerals. They lack chloroplasts and have reddish walls.

FIGURE 6:The leaves of many mosses curl and press against the stem as they dry, forming capillary channels that conduct water (Doug Sokell/Visuals Unlimited).
Development. Growth of the gametophore begins when a spore germinates and sends out a long, slender chlorophyllous cell. This cell undergoes mitosis and produces a branched system of similar cells; the entire network is a protonema (pl.: protonemata) (Fig. 7). A protonema superficially resembles a filamentous green alga but can be distinguished by its numerous small chloroplasts in each cell. Algal cells have only one or two large chloroplasts. Also, the cross walls of the protonema are placed at an angle to the side walls, not perpendicularly as in a green alga. Rhizoids grow from the underside of the protonema. Eventually nodules of small cytoplasmic cells form on the protonema. This buds organize an apical cell, then grow upright as a stem with leaves—the gametophore.
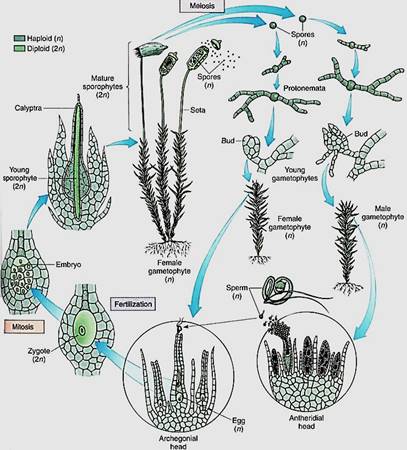
FIGURE 7:Life cycle of a moss.
Protonemata are perennial and can grow extensively, producing many buds. The filamentous cells usually break when mosses are collected, so a tuft of gametophores may appear to be independent plants when in fact they have all arisen from a single protonema.
Reproduction. The gametophore at some point produces gametangia. All mosses are oogamous; that is, every species has small biflagellate sperm cells and large nonmotile egg cells. Sperms are produced in microgametangia called antheridia, which consist of a short stalk, an outermost layer of sterile cells, and an inner mass of cells that differentiate into spam cells . Eggs occur in megagametangia called archegonia: Each is shaped like a vase with a long neck . The neck is hollow at maturity, and the single egg is located at the base.
Antheridia and archegonia occur on the same gametophore in bisexual species (Funaria, Pottia), whereas other species have both male and female gametophores (Barbula, Polytrichum, Rhacomitrium). Depending on the species, gametangia may occur mixed with leaves along the gametophore stem or may be clustered at the stem tip, surrounded by a special cup-shaped whorl of leaves (Fig. 8). This apical grouping appears to be especially advantageous because raindrops fall directly into the cup, splashing sperms out and carrying them as far as 50 cm. Without this mechanism, sperms must swim to archegonia.
When sperm cells are mature, the antheridium breaks open and liberates sperm cells either by contracting the sterile outer cells or by accumulating liquid below the sperms and pushing them out. Antheridia open when the gametophore is wet and a film of water is present through which sperms can swim (Fig. 9). The microgametes move to the archegonia either by swimming alone or by being splashed and then swimming. Secretion of sucrose from the archegonia guides sperms toward the archegonia and then down the neck to the egg, where one sperm cell effects fertilization.

FIGURE 8: (a) In this moss, antheridia are clustered together at the gametophore apex; in other species, they might be located along the stem. (E. R Degginger) (b) The phenomenon of dwarf males is common in many plants, animals, and algae. Each sperm cell is much smaller and less expensive to produce than each egg, but usually many sperm cells are not carried to eggs, so a large number of spam cells must be produced. If sperm delivery can be ensured, the number produced can be decreased, and even a small, dwarf male can produce adequate numbers. Deliver)' can be assured if the male gametophyte grows on the female as an epiphyte.

FIGURE 9: (a) The organelles of a young sperm cell have rather typical shapes. (Blue, nucleus; green, chloroplast; red, flagella and basal bodies; brown, mitochondrion; yellow, microtubules that anchor the flagella). (b and c) During differentiation, the organelles become modified, resulting in an elaborate spiral-shaped sperm cell at maturity (d). Hyaloplasm and cell membrane are not shown. (a, b, and d, Courtesy of Karen Renzaglia and Fred Alsop HI; c, courtesy of Douglas Bernhard and K. Renzaglia. East Tennessee State University)
THE SPOROPHYTE GENERATION
In all embryophytes, the megagamete and subsequently the zygote are retained by the gametophyte. However, unlike those of mosses, megagametophytes of vascular plants are small and offer little nourishment or protection to the embryonic sporophytes. Megagametophytes of flowering plants typically have only six cells other than the egg, and both synergids and antipodals often degenerate just after the egg is fertilized (undergoes syngamy). All nutrition for the zygote is supplied by the grandparent generation, the sporophyte, by way of endosperm formation. In contrast, moss gametophytes are both large and very efficient at photosynthesis, and they support the sporophyte throughout its entire life The moss sporophyte is never an independent, free-living plant .
The zygote of a moss undergoes a transverse division, and the basal cell, located at the bottom of the archegonium, develops into a small, bulbous tissue called the foot. He foot is the interface with the gametophore, from which it absorbs sugars, minerals, and water: Its cells are transfer cells in many species. The upper cell grows by cell division and expansion into a simple apical sporangium called the capsule, consisting of an outer layer of sterile cells and an inner column of sterile cells (the columella). A ring of sporogenous cells undergoes meiosis, producing haploid spores (Fig. 10). Between the foot and the sporangium is a narrow stalk, the seta (pl.: setae). All moss sporophytes have this basic, simple structure; none is ever branched or has leaves, bracts, or buds of any kind.

FIGURE 10:Aspects of a moss sporophyte. (a) The foot is embedded in the remnants of the archegonium. (b) External view of the capsule (sporangium) with the operculum still in place covering the teeth(c) Median section view showing the columella and spores. (d) External view after the operculum has fallen off. Spores can be shed when the peristome teeth bend back. (e) Scanning electron micrograph view of peristome teeth. (G. Shih and R Kessel/Visuals Unlimited)
Although morphologically simple, the sporophyte is relatively complex structurally. It is smaller than the gametophyte but more dense, being many cells in diameter, even in the seta and foot regions. The sporophyte has a true epidermis with stomata at least on the base of the sporangium . Below the epidermis the seta contains long, thick-walled cells and often leptoids and hydroids. Dehiscence of the sporangium is more elaborate than the opening of the gametangia: The apex of the sporangium differentiates as a caplike lid, the operculum, which separates from the rest of the sporangium as cells are torn apart. Cell breakage is elaborate and precise, resulting in one or two rows of beautiful, exquisitely complex teeth, called peristome teeth (Fig. 10). The teeth respond to humidity, bending outward and opening the sporangium when the air is dry and bending inward and trapping the spores when the air is humid. Spores are released when they are light, dry, non-sticky, and easily carried by air currents (Fig. 11).
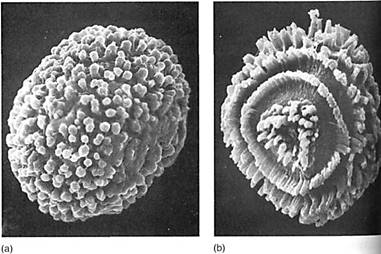
FIGURE 11:The transition to land required the evolution of spores that could resist desiccation. Little is known about the chemicals that provide waterproofing. Like pollen, moss spores show many types of surface pattern when studied by scanning electron microscopy, (a) Acaulon rufescens (X 600). (b) Bruchia drummondi (X 600). (Courtesy of D. M. ]. Mueller, Texas A and M University)
In addition to the operculum, the apex of the sporangium in many species is covered by a calyptra, a layer of cells derived from the neck of the archegonium. As the embryo begins to grow, neck cells also proliferate. They keep pace with sporophyte growth at first but later grow more slowly and are torn away from the gametophore. The calyptra is important for proper development; if it is prematurely cut or removed surgically, the seta elongates but a sporangium does not form.
Virtually all mosses are homosporous: All spores are the same size and appear to be identical. A few species of Macromitrium and Schlotheimia produce two types of spores in each capsule. The larger spores develop into large gametophytes with archegonia, and the smaller spores grow into dwarf males that live epiphytically on the females.
METABOLISM AND ECOLOGY
Small size and lack of conducting tissues are two critical factors in the metabolism and ecology of mosses. Vascular plants tend to be larger than nonvascular plants, and their bulk protects them from short-term fluctuations in air humidity and moisture availability. Should the air become dry for several hours, most leaves and stems would not die, even if they were not receiving water from the roots by means of xylem. Very large plants, such as trees and succulents, can withstand dry conditions for weeks, months, or even years before losing a fatal amount of water. Mosses, however, have no such mechanisms to absorb and store water. Leaves on the gametophore have only a thin, incomplete cuticle, and typically at least the undersides are not cutinized. If exposed to dry air for even a few minutes, the plants dry out. Without vascular tissue, stems and leaves can become desiccated, even while the rhizoids are in contact with moist soil or tree bark.
Several mechanisms compensate for the inability of mosses to retain water. Many species grow in permanently moist microhabitats such as rainforests, cloud forests, and the spray zones near waterfalls (Fig. 12a). In such situations, the plants can become quite large because water is available to and absorbed by the entire plant suriace, so water-conducting tissues are completely unnecessary. Even habitats that are not constantly humid may have microhabitats that are continually moist. The foot of a rock cliff is usually damp because dew and mist concentrate there as runoff from the cliff. Shallow depressions in rock or soil retain moisture and are protected from drying winds. These sites are very small, but so are the mosses that live within them.
Other mosses compensate for their inability to retain water by being tolerant of desiccation. That is, drying does not damage them as it does most vascular plants and algae. Like lichens, many mosses can lose much of their water rather rapidly without dying or even being injured. As long as about 30% of their weight is water, they remain dormant but alive. If rain falls or dew forms, water is absorbed rapidly, and within a few minutes respiration and photosynthesis are occurring at normal levels.

FIGURE 12: (a) Most mosses live in habitats that are permanently wet, never drying or subjecting the mosses to wafer stress. (Sydney Karp, PHOTO/NATS) (b) Sphagnum moss grows at the edges of streams or ponds where there is little water movement. The plants become large and tangled, creating a mat that is solid enough to support the weight of an animal. These are known as quaking bogs, because they quake and undulate with every step. Given enough time, seeds of vascular plants germinate, and leaves and debris blow onto the bog and decompose, slowly converting it to a rich soil. (G. Bamgarter, PHOTO/NATS)
The alternation between dry and turgid states varies with the environment. In mild temperate regions, mosses may remain moist and metabolically active except for certain times during summer months. In deserts and dry range lands, on the other hand, mosses may be turgid primarily during winter. From spring to autumn, the mosses are dry and inactive every day, perhaps receiving enough dew on certain nights to be metabolically active for a few hours in the early morning. In harsh conditions, gametophores grow crowded together, forming an extremely dense clump. The close packing prevents air movement and helps retain transpired water molecules, and dew or rain that touches the top of the tuft is conducted downward by capillary action.
Desiccated mosses are remarkably resistant to high or low temperature and to intense ultraviolet (UV) light. A common moss, Tortula ruralis, can be frozen safely in liquid nitrogen (—196°C) when dry, and can also withstand brief periods at +100°C. Resistance to UV light is important for species that grow in full sun on the surface of rocks or at high altitudes on mountains, where the atmosphere is too thin to block UV radiation.
Many moss species thrive at low temperatures near or even below 0°C This is adaptive in habitats where the larger vascular plants die or abscise their leaves in winter. Leaf fall permits much more sunlight to reach the soil surface or tree trunks where the mosses are growing. Snow or winter rains provide abundant moisture. Mosses that can carry out metabolism at cold winter temperatures can take advantage of the plentiful light and moisture.
Certain mosses can grow on hard, impervious surfaces because they have no roots that must penetrate the substrate. Consequently, mosses can be found on bare rock (Andreaea, Hedwigia), brick or mortar (Tortula muralis), and bark as well as soil. At the other extreme, Sphagnum mosses live on the surface of quiet water at the edges of lakes and ponds . Their leaves and stems arc buoyant, and such thick mats can be built up that a person can walk across the floating colony if enough care is taken.
In almost all environments, mosses are important in the later establishment of other species, particularly vascular plants. Like lichens, they can colonize a bare surface and dissolve the rock with acids that leak from their cells. Leaves and stems of gametophores catch and hold dust particles, creating small pockets of soil in which spores and seeds of vascular plants can become established. A similar process occurs even on the ground; in forests or grasslands, the moss layer on the soil may be thick enough to act as a moist, airy seed bed. It may hold water better than the soil and thus improve the microhabitat for seedlings.



|
|
|
|
دخلت غرفة فنسيت ماذا تريد من داخلها.. خبير يفسر الحالة
|
|
|
|
|
|
|
ثورة طبية.. ابتكار أصغر جهاز لتنظيم ضربات القلب في العالم
|
|
|
|
|
|
|
وفد العتبة الكاظمية المقدسة يُلبّي دعوة حضور حفل افتتاح مهرجان السفير الثقافي بنسخته الرابعة عشر
|
|
|