آخر المواضيع المضافة

 النبات
الحيوان
الأحياء المجهرية
علم الأمراض
التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحياتية النانوية
علم الأجنة
الأحياء الجزيئي
علم وظائف الأعضاء
الغدد
المضادات الحيوية
النبات
الحيوان
الأحياء المجهرية
علم الأمراض
التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحياتية النانوية
علم الأجنة
الأحياء الجزيئي
علم وظائف الأعضاء
الغدد
المضادات الحيوية| Environmental and Internal Factors |


|
|
Read More
Date: 18-10-2016
Date: 9-11-2016
Date: 30-10-2016
|
Environmental and Internal Factors
A plant's photosynthesis is affected by its environment in many ways.
LIGHT
From a plant's viewpoint, light has three important properties: (1) quality, (2) quantity, and (3) duration.
Quality of sunlight refers to the colors or wavelengths that it contains. Sunlight is pure while because it contains the entire visible spectrum. During sunset and sunrise, sunlight passes tangentially through the atmosphere and a large percentage of the blue light is deflected upward; consequently, light at ground level is enriched in red, which is easily visible. This period of red-enriched light lasts only a few minutes and probably has little effect on photosynthesis. At noon, sunlight passes nearly vertically through the atmosphere, more blue light is transmitted, and even though the blueness of the sky suggests that all of the reds, greens, and yellows have been blocked, in fact enough of all of these wavelengths penetrate to the Earth's surface to allow efficient photosynthesis. This is true of plants in deserts, grasslands, and the top layer, the canopy, of a forest. But herbs and shrubs that grow near soil level in a forest are understory plants, and the light they receive has already passed through the leaves of the canopy (Fig.1). As light penetrates those leaves, red and blue are absorbed by chlorophyll, so the dim light received by understory plants is especially depleted in these critical wavelengths. It is selectively advantageous for them to have extra amounts of accessory pigments so that they can gather the wavelengths available and pass the energy on to chlorophyll a. Similarly, algae that grow near the surface of lakes or oceans receive complete light, but water absorbs red and violet. Algae at deep regions receive mostly green and blue light and must have special accessory pigments capable of absorbing these wavelengths efficiently.
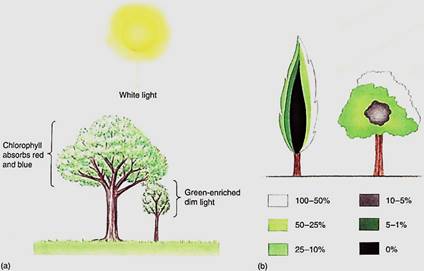
FIGURE 1 :(a) Plants growing in the shade of other plants receive not only dim light, but light depleted of BC and blue. (b) Not all leaves of a tree receive equal amounts of light; this self-shading is more severe in latitudes farther away fern the equator. Colors indicate the percentage of full sunlight that reaches the various parts of a tree.
Quantity of light, which refers to light intensity or brightness, is affected by several factors. More light is available for photosynthesis on a clear than on a cloudy day; under y plants receive dim light; lower branches and branches on the shaded side of a plant
receive less light (Fig. 1b). Plants growing in the shadow of mountains or in deep canyons receive much less light than plants that grow on slopes that face the sun. Plains growing near the equator receive intense light because the sun is always more or less directly overhead at noon, whereas plants near the poles receive very little light. Even during the summer the sun is low at noon and light is scattered by the atmosphere.
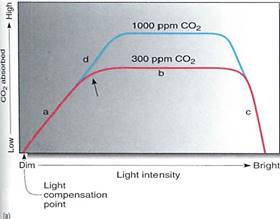

FIGURE 2 :(a) Light and photosynthesis; details are given in text. The unt ppm is parts per million; for 300 ppm, in 1 million liters of air, 300 liters are carbon dioxide. (b) These plants have aburdant sunlight. They could probably photosynthesize me rapidly if more carbon dioxide were present.
Think about how intensity of sunlight varies during the day and affects photosynthe¬sis. Examine the solid line labeled "300 ppm CO2" in Figure 2. This was derived im many experiments in which the rate of photosynthesis was measured for plants gown under different intensities of light, but all with 300 ppm carbon dioxide in the air. Near the left side at point a, those plants that received dim light absorbed little carbon cficxide, whereas those grown in brighter light absorbed more carbon dioxide. Under these romd levels of carbon dioxide, light is the limiting factor. Photosynthesis is slow on dull, aerat days but faster on brighter days. At point b in the graph, plants that received more light dd not photosynthesize faster than those that received slightly less. Where the curve turns flat, there was enough light to saturate the process. In these conditions, the limitation was kk of carbon dioxide; in the experiments in which more carbon dioxide was available (tie line, 1000 ppm CO2), photosynthesis went faster. Thus at point b, carbon dioxide was he rate-limiting factor. At point c, light was so intense that it damaged the plant by overheating it and bleaching the pigments.
At point a, if the lack of light prevents photosynthesis from proceeding faster, there must be adequate amounts of carbon dioxide. As soon as ATP and NADPH are produced, they move to the stroma and are used by the waiting enzymes and carbon dioxide; then ADP and NADP+ diffuse back to the thylakoid membranes and wait for another quantum. Conversely, at point b, where light is bright and the low concentration of carbon dbxde is rate limiting, there is so much light that as quickly as ADP and NADP+ come to the thylakoids, they are reprocessed immediately into ATP and NADPH. These then move to the stroma, where they must wait for a carbon dioxide molecule.
To the left of the light compensation point, it appears that there was no phoosyn- thesis, even though some dim light was provided. The problem actually lies with the technology of measuring photosynthesis. Either the amount of carbon dioxide absorbed or the amount of oxygen released must be measured. But both of these gases are involved in respiration as well as photosynthesis. To the left of the light compensation point, photo- synfibesis was absorbing carbon dioxide more slowly than respiration was releasing it, so it appears as though no photosynthesis occurred. The same problem arises when we try to neaare respiration: Photosynthesis distorts the measurement, but then we can turn off the lights and stop photosynthesis. The light compensation point is the level of light at which photosynthesis matches respiration. Plants that are grown for a long time in conditions b±w the light compensation point respire faster than they photosynthesize; they gradually comme their reserve carbohydrates and fats and starve to death. For plants grown in light brighter than the light compensation point, photosynthesis exceeds respiration and the extra sugar can be used for growth and reproduction.
In the brightest of environments, the air is so clear that sunlight is frequently too inte during summer months. Protective adaptations are necessary, and in many species mechanisms have evolved that provide shade. A common method is the production of a thick layer of dead trichomes, plant hairs (Fig. 3). A heavy coating of wax can also reflect, light, and cutin is especially good at absorbing the more harmful short wavelengths. Part of the value of carotenoids and other accessory pigments is that they shade the chloro-phyll, absorbing some of the most damaging wavelengths.
The intensity of light, the actual number of quanta that strike a given area per unit time (eg, that strike 1 cm2/s) is greatest at noon in midsummer when the sun is most directly overhead and is less whenever the sun is lower—morning, evening, and winter. Light may be teo intense at midday but optimal when the sun is lower; some species (iris, eucalyptus) have adapted to this by means of vertical leaves. The lamina face is exposed fully only in mornings and afternoons, but at noon only the leaf edge is exposed. In other plants, leaves orient vertically automatically when stressed—they wilt and hang down.
Understory plants of forests are adapted to low light. If a roadway is cut into a forest and plants adjacent to the cut are suddenly exposed to full sunlight, the shock may cause them to wilt and die. The same phenomenon occurs when trees are blown down during sdm$ floods, or avalanches and the surrounding plants are exposed.

FIGURE 3 :(a) While young, the leaves of dusty miller are completely obscured by trichomes, protecting the leaf from strong sunlight and insects. (b) These cacti (Epithelantha) live in environments where sunlight is extremely intense; their spines are so abundant and closely spaced that they shade the stem and prevent the chlorophyll from being damaged. (Big Bend National Park).
Duration of light refers to the number of hours per day that sunlight is available. At the equator, days are 12 hours long throughout the year. Farther north or south, days become longer in summer; maximum length occurs near the poles, where the day is 24 hours long in midsummer and only night occurs in midwinter. In middle latitudes, winter days are short and sunlight is weak because the sun is so low in the sky. Under these conditions, even evergreen plants are unable to undergo very much photosynthesis. However, because temperatures are low, the plants are growing little and have a low rate of respiration. Even deciduous trees and biennials can survive by means of stored nutrients.
During summer, days are longer and light is brighter because the sun is higher in the sky. The amount of energy obtained by photosynthesis easily exceeds the amount consumed by respiration and growth. In many plants, longer days cause greater amounts of photosynthesis, but in others, chloroplasts become so full of starch that photosynthesis stops, even though light is present. At night, starch is converted to sugar, which is fan transported out of chloroplasts and can be used for growth or stored in amyloplasts in tubers, corms, or other such organs. By morning, leaf chloroplasts can resume photosynthesis.
LEAF STRUCTURE
heaf structure of most temperate and tropical plants is quite standard: palisade parenchyma above and spongy mesophyll below. This structure is excellent for absorbing carbon dioxide but inefficient for conserving water. If plants of hot, dry habitats had this leaf architecture, they would have to keep their stomata closed so much of the time that they would starve. Instead, their leaf cells are frequently packed together without intercellular spaces. Water loss is reduced because the small internal surface area retards water evaporation. But with so little surface area, it is difficult to dissolve carbon dioxide from the air into the cytoplasm. This slows photosynthesis, but apparently this trade-off, slow growth versus water conservation, is selectively advantageous.
Another method of minimizing water loss while maintaining photosynthesis is to reduce external surface by means of cylindrical leaves. Water movement from interior air spaces to exterior air is minimized because so few stomata are present (Fig. 4). Photosynthesis is reduced because absorption of carbon dioxide is slowed.

FIGURE 10.24:These living stone plants (Lithops) of African deserts conserve water in several ways. They have only two leaves at a time; when two new ones form, the old two die. Not enough water is available for four leaves. The leaves are fleshy and pressed together, such that they form a cylinder with minimal surface area through which water can be lost.
WATER
The amount of water available greatly affects photosynthesis. Most plants keep their stomala open during the clay, permitting the entry of carbon dioxide, but water is inevitably lost. At night, carbon dioxide cannot be used, and stomata are closed, retaining water within the plant. If the soil becomes dry and water is not readily available, a plant keeps its stomata closed even during the day, and carbon dioxide cannot enter (Fig.5). The small amount of carbon dioxide produced by the respiration of the leaves can be reused photosynthetically, but this is a minor amount. In addition to numerous structural modifications that conserve water, metabolic adaptations also exist; two of the most important are C4 metabolism and crassulacean acid metabolism.
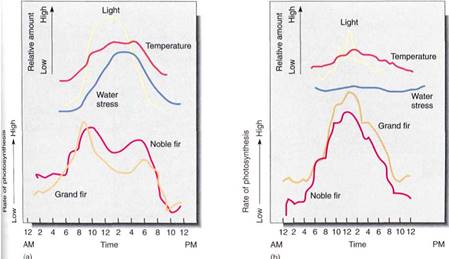
FIGURE 5: (a) Tight intensity exceeded the light compensation point for these trees just after 6 AM, and photosynthesis increased rapidly. However, by early afternoon, photosynthesis dropped even thigh light and temperature were adequate; the problem was lack of water (water stress), and probably stomata had begun to close around noon. (b) On a cloudy day, water stress never became a problem and stomata remained open. Even though there was less total light than in (a), these was more photosynthesis for the day.
C4 METABOLISM
An important factor for plants is the amount of water lost for each molecule of carbon dioxide absorbed. Ideally, this ratio is low. Carbon dioxide diffuses into a leaf faster if its concentration in air is higher or if its concentration inside leaf protoplasm is lower. Plants can do nothing about the carbon dioxide level in the external air, and many also have poor control over the protoplasmic concentration of carbon dioxide. RuBP carboxylase has a low affinity for carbon dioxide; as carbon dioxide concentration drops, enzyme-substrate binding slows. Even while carbon dioxide is still rather abundant in protoplasm, the enzyme is only rarely picking it up. With this relatively high concentration, carbon dioxide diffusion into the leaf is slow whereas water loss may be high.
RuBP carboxylase occasionally binds to oxygen instead of carbon dioxide, acting as an oxygenase and producing one molecule of 3-phosphoglycerate and one of phosphoglycolate. This latter molecule is transported from the chloroplast to peroxisomes and mitochondria, where some of it is converted to the useful amino acids glycine and serine. But much of the phosphoglycolate is broken down to two molecules of carbon dioxide (Fig.6). The breakdown is photorespiration, an energy-wasting process. The energy and reducing power used to produce the two reduced carbons of phosphoglycolate are completely lost. Photorespiration is extremely exergonic but is not used to provide power to endergonic reactions. Because phosphoglycolate may be toxic, photorespiration protects the plant whenever RuBP carboxylase picks up oxygen. However, it is an expensive defense because up to 30% of all ATP and NADPH produced by the chloroplast can be immediately lost by photorespiration.
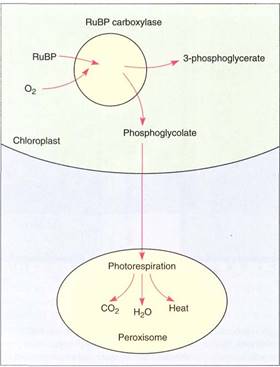
FIGURE 6 :If RuBP carboxylase puts oxygen onto RuBP, one of the products is phosphoglycolate, which is transported to a peroxisome and broken down during photorespiration. All the energy that was present in the phosphoglycolate is wasted. Mitochondria are also involved.
Apparently RuBP carboxylase cannot be significantly improved. At the time when RuBP carboxylase originated by evolution, virtually no free oxygen was present in the atmosphere, and carbon dioxide levels were high . RuBP carboxylase has existed for billions of years, and virtually no structural mutations have survived, so they must not have produced superior versions of the enzyme. An alternative is to improve its working conditions. RuBP carboxylase should be compartmentalized in a site where carbon dioxide concentration is high and oxygen concentration is low. This has evolved in some plant groups and is known as C4 metabolism or C4 photosynthesis. Basically, C4 metabolism is a mechanism by which carbon dioxide is absorbed, transported through, and concentrated in a leaf, whereas oxygen is kept away from RuBP carboxylase
C4 metabolism occurs in leaves with Kranz anatomy (Fig. 7). In such leaves, mesophyll is not distributed as palisade and spongy parenchyma, but rather each vascular bundle has a prominent chlorophyllous sheath of cells and around the sheath are mesophyll cells. Mesophyll cells contain the enzyme PEP (phosphoenolpyruvate) carboxylase, which has a very high affinity for carbon dioxide. Unlike RuBP carboxylase, as the carbon dioxide concentration drops lower and lower, PEP carboxylase continues binding to it rapidly and firmly. Carbon dioxide concentrations inside the leaf are kept very low and carbon dioxide diffuses inward rapidly whenever stomata are open. The ratio of water lost to carbon dioxide absorbed is favorably low. Also, PEP carboxylase has a high specificity for carbon dioxide; it never picks up oxygen. PEP carboxylase is an ideal enzyme except that it does not perform the critical reaction that results in 3-phosphoglycerate. Despite its shortcomings, RuBP carboxylase is still the only enzyme that carries out the necessary reaction.

FIGURE 7 :(a) In leaves with Kranz anatomy, such as this sugarcane, the bundle sheath around all veins is prominent and rich in chloroplasts. The mesophyll cells are arranged in a sheath around the bundle sheath (X 150). (b) Knowing the reactions of C4 metabolism is not sufficient for understanding it; only by realizing that some reactions occur in separate compartments does it become logical.
PEP carboxylase adds carbon dioxide to PEP, producing oxaloacetate, which has four carbons; hence the name C4 metabolism. The oxaloacetate is reduced to malate by a molecule of NADPH, and further reactions may occur, depending on the species. Malate from throughout the mesophyll moves into the bundle sheath and breaks down into pyruvate by releasing carbon dioxide (Fig. 7b). This reaction is powerful enough to drive the formation of a new molecule of NADPH, so the process results in the transport by malate of both carbon dioxide and reducing power. Because all the malate from a large volume of mesophyll decarboxylates in the small volume of the bundle sheath, carbon dioxide concentration in the sheath is very high (Fig. 8). Also, because NADPH is synthesized by this unusual method in bundle sheath cells, the bundle sheath chloroplasts primarily carry out cyclic electron transport, pumping protons and making ATP. Without noncyclic electron transport, there is no breakdown of water or production of oxygen.
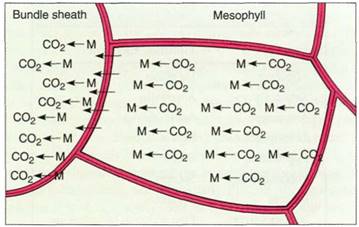
FGURE 8:Mesophyl cells are much larger than bundle sheath cells, so transport of carbon dioxide from mesophyll to sheath automatically results in a higher concentration in the sheath.
RuBP carboxylase, located exclusively in the bundle sheath chloroplasts, is in ideal conditions—high carbon dioxide concentration with low or no oxygen—and carries out efficient C3 stroma reactions with low production of phosphoglycolate. C4 plants have little photorespiration (Table1 ).

Take any other carrier, malate must be shuttled back to its recharge site, the mesophyll. In the bundle sheath it is converted to pyruvate by the release of carbon dioxide; pyruvate moves back to the mesophyll and receives a phosphate group from ATP, which converts it to PEP.
The selective advantage of C4 metabolism depends on the environment. Photorespiration increases with temperature, so it is more of a problem in hot climates. Under warm, dry conditions, C4 metabolism has a strong selective advantage over C3 metabolism: Much less water is lost during carbon dioxide absorption. Also, abundant light is available to generate the extra ATP needed to convert pyruvate to PEP. The ATP used to make HP means that C4 metabolism is not free; under cool conditions, photorespiration may be sow enough that it loses less energy than C4 metabolism. Also, many cool habitats are also moist, so water conservation by stomatal closure is not as critical. The critical temperature above which C4 metabolism is more advantageous selectively than C3 metabolism varies among species but averages around 25°C.
C4 metabolism and Kranz anatomy have evolved several times; most C4 species are monocots of hot climates such as corn, sugarcane, sorghum, and several other grasses, but a considerable number of dicots, also from warm, dry regions, are also C4 species (Table 2).




|
|
|
|
دخلت غرفة فنسيت ماذا تريد من داخلها.. خبير يفسر الحالة
|
|
|
|
|
|
|
ثورة طبية.. ابتكار أصغر جهاز لتنظيم ضربات القلب في العالم
|
|
|
|
|
|
|
العتبة العباسية المقدسة تستعد لإطلاق الحفل المركزي لتخرج طلبة الجامعات العراقية
|
|
|