آخر المواضيع المضافة

 الفيزياء الكلاسيكية
الكهربائية والمغناطيسية
علم البصريات
الفيزياء الحديثة
النظرية النسبية
الفيزياء النووية
فيزياء الحالة الصلبة
الليزر
علم الفلك
المجموعة الشمسية
الطاقة البديلة
الفيزياء والعلوم الأخرى
مواضيع عامة في الفيزياء
الفيزياء الكلاسيكية
الكهربائية والمغناطيسية
علم البصريات
الفيزياء الحديثة
النظرية النسبية
الفيزياء النووية
فيزياء الحالة الصلبة
الليزر
علم الفلك
المجموعة الشمسية
الطاقة البديلة
الفيزياء والعلوم الأخرى
مواضيع عامة في الفيزياء| Neurology of vision |


|
|
أقرأ أيضاً
التاريخ: 2023-04-12
التاريخ: 2024-04-06
التاريخ: 2023-11-19
التاريخ: 2023-12-12
|
One of the main points of our subject is the interconnection of information from one part of the eye to the other. Let us consider the compound eye of the horseshoe crab, on which considerable experimentation has been done. First of all, we must appreciate what kind of information can come along nerves. A nerve carries a kind of disturbance which has an electrical effect that is easy to detect, a kind of wavelike disturbance which runs down the nerve and produces an effect at the other end: a long piece of the nerve cell, called the axon, carries the information along, and a certain kind of impulse, called a “spike,” goes along if it is excited at one end. When one spike goes down the nerve, another cannot immediately follow. All the spikes are of the same size, so it is not that we get higher spikes when the thing is more strongly excited, but that we get more spikes per second. The size of the spike is determined by the fiber. It is important to appreciate this in order to see what happens next.

Fig. 36–11. The compound eye of the horseshoe crab. (a) Normal view. (b) Cross section. (Figures 36-7, 11, 12, 13 reprinted with permission from Goldsmith, Sensory Communication, W. A. Rosenblith, ed. Copyright 1961, Massachusetts Institute of Technology.)
Figure 36–11(a) shows the compound eye of the horseshoe crab; it is not very much of an eye, it has only about a thousand ommatidia. Figure 36–11(b) is a cross section through the system; one can see the ommatidia, with the nerve fibers that run out of them and go into the brain. But note that even in a horseshoe crab there are little interconnections. They are much less elaborate than in the human eye, and it gives us a chance to study a simpler example.
Let us now look at the experiments which have been done by putting fine electrodes into the optic nerve of the horseshoe crab, and shining light on only one of the ommatidia, which is easy to do with lenses. If we turn a light on at some instant t0, and measure the electrical pulses that come out, we find that there is a slight delay and then a rapid series of discharges which gradually slow down to a uniform rate, as shown in Fig. 36–12(a). When the light goes out, the discharge stops. Now it is very interesting that if, while our amplifier is connected to this same nerve fiber, we shine light on a different ommatidium nothing happens; no signal.
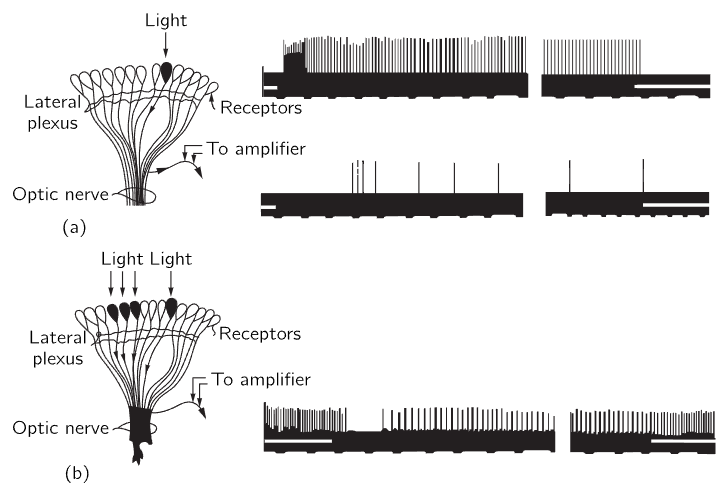
Fig. 36–12. The response to light of the nerve fibers of the eye of the horseshoe crab.
Now we do another experiment: we shine the light on the original ommatidium and get the same response, but if we now turn light on another one nearby as well, the pulses are interrupted briefly and then run at a much lower rate (Fig. 36–12b). The rate of one is inhibited by the impulses which are coming out of the other! In other words, each nerve fiber carries the information from one ommatidium, but the amount that it carries is inhibited by the signals from the others. So, for example, if the whole eye is more or less uniformly illuminated, the information coming from any one ommatidium will be relatively weak, because it is inhibited by so many. In fact, the inhibition is additive—if we shine light on several nearby ommatidia the inhibition is very great. The inhibition is greater when the ommatidia are closer, and if the ommatidia are far enough away from one another, inhibition is practically zero. So, it is additive and depends on the distance; here is a first example of information from different parts of the eye being combined in the eye itself. We can see, perhaps, if we think about it awhile, that this is a device to enhance contrast at the edges of objects, because if a part of the scene is light and a part is black, then the ommatidia in the lighted area give impulses that are inhibited by all the other light in the neighborhood, so it is relatively weak. On the other hand, an ommatidium at the boundary which is given a “white” impulse is also inhibited by others in the neighborhood, but there are not as many of them, since some are black; the net signal is therefore stronger. The result would be a curve, something like that of Fig. 36–13. The crab will see an enhancement of the contour.
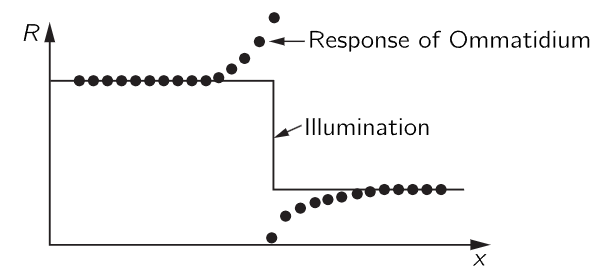
Fig. 36–13. The net response of horseshoe crab ommatidia near a sharp change in illumination.
The fact that there is an enhancement of contours has long been known; in fact, it is a remarkable thing that has been commented on by psychologists many times. In order to draw an object, we have only to draw its outline. How used we are to looking at pictures that have only the outline! What is the outline? The outline is only the edge difference between light and dark or one color and another. It is not something definite. It is not, believe it or not, that every object has a line around it! There is no such line. It is only in our own psychological makeup that there is a line; we are beginning to understand the reasons why the “line” is enough of a clue to get the whole thing. Presumably our own eye works in some similar manner—much more complicated, but similar.
Finally, we shall briefly describe the more elaborate work, the beautiful, advanced work that has been done on the frog. Doing a corresponding experiment on a frog, by putting very fine, beautifully built needlelike probes into the optic nerve of a frog, one can obtain the signals that are going along one particular axon and, just as in the case of the horseshoe crab, we find that the information does not depend on just one spot in the eye, but is a sum of information over several spots.
Table 36–1
Types of response in optic nerve fibers of a frog
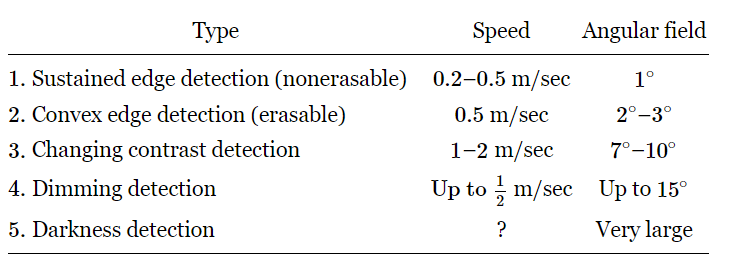
The most recent picture of the operation of the frog’s eye is the following. One can find four different kinds of optic nerve fibers, in the sense that there are four different kinds of responses. These experiments were not done by shining on-and-off impulses of light, because that is not what a frog sees. A frog just sits there and his eyes never move, unless the lily pad is flopping back and forth, and in that case his eyes wobble just right so that the image stays put. He does not turn his eyes. If anything moves in his field of vision, like a little bug (he has to be able to see something small moving in the fixed background), it turns out that there are four different kinds of fibers which discharge, whose properties are summarized in Table 36–1. Sustained edge detection, nonerasable, means that if we bring an object with an edge into the field of view of the frog, then there are a lot of impulses in this particular fiber while the object is moving, but they die down to a sustained impulse that continues as long as the edge is there, even if it is standing still. If we turn out the light, the impulses stop. If we turn it on again while the edge is still in view, they start again. They are not erasable. Another kind of fiber is very similar, except that if the edge is straight, it does not work. It must be a convex edge with dark behind it! How complicated must be the system of interconnections in the retina of the eye of the frog in order for it to understand that a convex surface has moved in! Furthermore, although this fiber does sustain somewhat, it does not sustain as long as the other, and if we turn out the light and turn it on again it does not build up again. It depends on the moving in of the convex surface. The eye sees it move in and remembers that it is there, but if we merely turn out the light for a moment, it simply forgets it and no longer sees it.
Another example is change-in-contrast detection. If there is an edge moving in or out there are pulses, but if the thing stands still there are no pulses at all.
Then there is a dimming detector. If the light intensity is going down it creates pulses, but if it stays down or stays up, the impulse stops; it only works while the light is dimming.
Then, finally, there are a few fibers which are dark detectors—a most amazing thing—they fire all the time! If we increase the light, they fire less rapidly, but all the time. If we decrease the light, they fire more rapidly, all the time. In the dark they fire like mad, perpetually saying, “It is dark! It is dark! It is dark!”
Now these responses seem to be rather complicated to classify, and we might wonder whether perhaps the experiments are being misinterpreted. But it is very interesting that these same classes are very clearly separated in the anatomy of the frog! By other measurements, after these responses had been classified (afterwards, that is what is important about this), it was discovered that the speed of the signals on the different fibers was not the same, so here was another, independent way to check which kind of a fiber we have found!
Another interesting question is from how big an area is one particular fiber making its calculations? The answer is different for the different classes.

Fig. 36–14. The tectum of a frog.
Figure 36–14 shows the surface of the so-called tectum of a frog, where the nerves come into the brain from the optic nerve. All the nerve fibers coming in from the optic nerve make connections in various layers of the tectum. This layered structure is analogous to the retina; that is partly why we know that the brain and retina are very similar. Now, by taking an electrode and moving it down in succession through the layers, we can find out which kinds of optic nerves end where, and the beautiful and wonderful result is that the different kinds of fibers end in different layers! The first ones end in number 1 type, the second in number 2, the threes and fives end in the same place, and deepest of all is number four. (What a coincidence, they got the numbers almost in the right order! No, that is why they numbered them that way, the first paper had the numbers in a different order!)
We may briefly summarize what we have just learned this way: There are three pigments, presumably. There may be many different kinds of receptor cells containing the three pigments in different proportions, but there are many cross connections which may permit additions and subtractions through addition and reinforcement in the nervous system. So before we really understand color vision, we will have to understand the final sensation. This subject is still an open one, but these researches with microelectrodes and so on will perhaps ultimately give us more information on how we see color.



|
|
|
|
للعاملين في الليل.. حيلة صحية تجنبكم خطر هذا النوع من العمل
|
|
|
|
|
|
|
"ناسا" تحتفي برائد الفضاء السوفياتي يوري غاغارين
|
|
|
|
|
|
|
ملاكات العتبة العباسية المقدسة تُنهي أعمال غسل حرم مرقد أبي الفضل العباس (عليه السلام) وفرشه
|
|
|