
آخر المواضيع المضافة

 النبات
الحيوان
الأحياء المجهرية
علم الأمراض
التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحياتية النانوية
علم الأجنة
الأحياء الجزيئي
علم وظائف الأعضاء
الغدد
المضادات الحيوية
النبات
الحيوان
الأحياء المجهرية
علم الأمراض
التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحياتية النانوية
علم الأجنة
الأحياء الجزيئي
علم وظائف الأعضاء
الغدد
المضادات الحيوية| Liver in Fasting |


|
|
Read More
Date: 20-12-2021
Date: 3-9-2021
Date: 17-11-2021
|
Liver in Fasting
The primary role of the liver in fasting is maintenance of blood glucose through the production of glucose (from glycogenolysis and gluconeogenesis) for glucose-requiring tissues and the synthesis and distribution of ketone bodies for use by other tissues. Therefore, hepatic metabolism is distinguished from peripheral (or extrahepatic) metabolism.
A. Carbohydrate metabolism
The liver first uses glycogen degradation and then gluconeogenesis to maintain blood glucose levels to sustain energy metabolism of the brain and other glucose-requiring tissues in the fasted state. [Note: Recall that the presence of glucose-6-phosphatase in the liver allows the production of free glucose both from glycogenolysis and from gluconeogenesis .]
1. Increased glycogenolysis: Figure 1 shows the sources of blood glucose after ingestion of 100 g of glucose. During the brief absorptive period, ingested glucose is the major source of blood glucose. Several hours later, blood glucose levels have declined sufficiently to cause increased secretion of glucagon and decreased secretion of insulin. The increased glucagon/insulin ratio causes a rapid mobilization of liver glycogen stores (which contain ~80 g of glycogen in the fed state) because of PKA-mediated phosphorylation (and activation) of glycogen phosphorylase kinase that phosphorylates (and activates) glycogen phosphorylase . Figure 1 shows that because liver glycogen is exhausted by 24 hours of fasting, hepatic glycogenolysis is a transient response to early fasting. Figure 2, , shows glycogen degradation as part of the overall metabolic response of the liver during fasting. [Note: Phosphorylation of glycogen synthase simultaneously inhibits glycogenesis.]

Figure 1: Sources of blood glucose after ingestion of 100 g of glucose. [Note: See Section B.2 for an explanation of the decline in gluconeogenesis.]

Figure 2: Major metabolic pathways in the liver during fasting. [Note: The numbers in circles, which appear both in the figure and in the corresponding citation in the text, indicate important metabolic pathways for carbohydrate or fat.] P = phosphate; CoA = coenzyme A; TCA = tricarboxylic acid; NADH = nicotinamide adenine dinucleotide.
2. Increased gluconeogenesis: The synthesis of glucose and its release into the circulation are vital hepatic functions during short- and long-term fasting (see Fig. 2, ). The carbon skeletons for gluconeogenesis are derived primarily from glucogenic amino acids and lactate from muscle and glycerol from adipose tissue. Gluconeogenesis, favored by activation of fructose 1,6-bisphosphatase (because of decreased availability of its inhibitor fructose 2,6-bisphosphate) and by induction of PEPCK by glucagon ), begins 4–6 hours after the last meal and becomes fully active as stores of liver glycogen are depleted (see Fig. 1). [Note: The decrease in fructose 2,6-bisphosphate simultaneously inhibits glycolysis at PFK-1 .]
B. Fat metabolism
1. Increased fatty acid oxidation: The oxidation of FA obtained from TAG hydrolysis in adipose tissue is the major source of energy in hepatic tissue in the fasted state (see Fig. 2, ). The fall in malonyl CoA because of phosphorylation (inactivation) of ACC by AMPK removes the brake on CPT-I, allowing β-oxidation to occur . FA oxidation generates NADH, flavin adenine dinucleotide (FADH2), and acetyl CoA. The NADH inhibits the TCA cycle and shifts OAA to malate. This results in acetyl CoA being available for ketogenesis. The acetyl CoA is also an allosteric activator of PC and an allosteric inhibitor of PDH, thereby favoring use of pyruvate in gluconeogenesis . [Note: Acetyl CoA cannot be used as a substrate for gluconeogenesis, in part because the PDH reaction is irreversible.] Oxidation of NADH and FADH2 coupled with oxidative phosphorylation supplies the energy required by the PC and PEPCK reactions of gluconeogenesis.
2. Increased ketogenesis: The liver is unique in being able to synthesize and release ketone bodies, primarily 3-hydroxybutyrate but also acetoacetate, for use as fuel by peripheral tissues but not by the liver itself because liver lacks thiophorase . Ketogenesis, which starts during the first days of fasting (Fig. 3), is favored when the concentration of acetyl CoA from FA oxidation exceeds the oxidative capacity of the TCA cycle. [Note: Ketogenesis releases CoA, insuring its availability for continued FA oxidation.] The availability of circulating water-soluble ketone bodies is important in fasting because they can be used for fuel by most tissues, including the brain, once their blood level is high enough.
Ketone body concentration in blood increases from ~50 μM to ~6 mM in fasting. This reduces the need for gluconeogenesis from amino acid carbon skeletons, thus preserving essential protein (see Fig. 1). Ketogenesis as part of the overall hepatic response to fasting is shown in Figure 2, . [Note: Ketone bodies are organic acids and, when present at high concentrations, can cause ketoacidosis.]
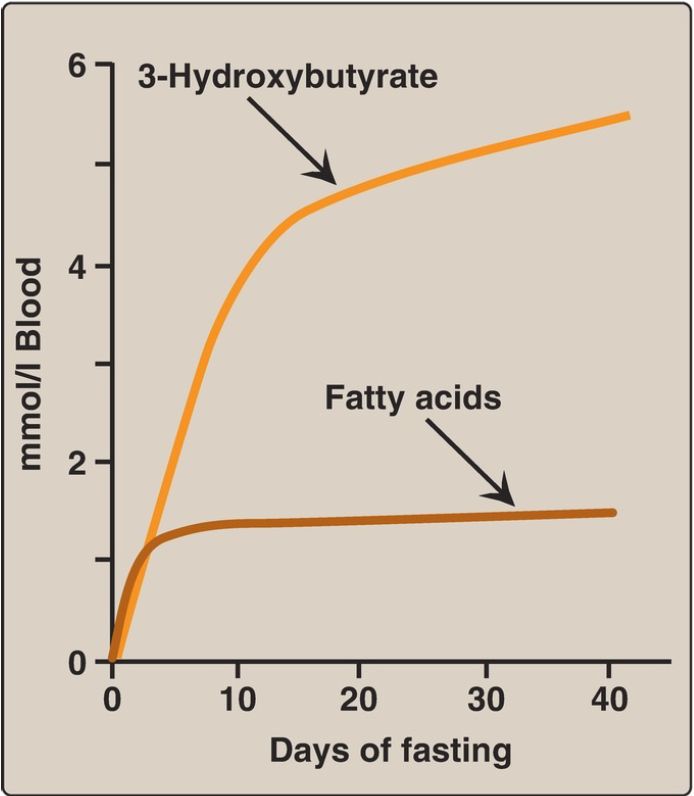
Figure 3: Concentrations of fatty acids and 3-hydroxybutyrate in the blood during fasting. [Note: 3-Hydroxybutyrate is made from the NADH-requiring reduction of acetoacetate.]



|
|
|
|
إدارة الغذاء والدواء الأميركية تقرّ عقارا جديدا للألزهايمر
|
|
|
|
|
|
|
شراء وقود الطائرات المستدام.. "الدفع" من جيب المسافر
|
|
|
|
|
|
|
بالصور: الامين العام للعتبة الحسينية المقدسة يجري جولة ميدانية للوقوف على آخر الاستعدادات الخاصة بمراسيم تبديل راية الإمام الحسين (ع)
|
|
|
















