آخر المواضيع المضافة

 النبات
الحيوان
الأحياء المجهرية
علم الأمراض
التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحياتية النانوية
علم الأجنة
الأحياء الجزيئي
علم وظائف الأعضاء
الغدد
المضادات الحيوية
النبات
الحيوان
الأحياء المجهرية
علم الأمراض
التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحياتية النانوية
علم الأجنة
الأحياء الجزيئي
علم وظائف الأعضاء
الغدد
المضادات الحيوية| CSR Involves AID and Elements of the NHEJ Pathway |


|
|
Read More
Date: 10-5-2016
Date: 21-4-2021
Date: 5-4-2021
|
CSR Involves AID and Elements of the NHEJ Pathway
KEY CONCEPTS
- Cross switch recombination (CSR) requires activation of intervening promoters (I promoters) that lie upstream of each of the two S regions involved in the recombination event and germline I -C transcription through the respective S regions.
- S regions contain highly repetitive 5′-AGCT-3′ motifs. 5′- AGCT-3′ repeats are the main targets of the CSR machinery and double-strand breaks (DSBs).
- Activation-induced deaminase (AID) mediates the first step (deoxycytidine deamination) in the series of events that lead to insertion of DSBs within S regions; the free ends of the DSBs are then religated through an NHEJlike reaction.
CSR initiates with transcription from the IH promoters of the CH regions that will be involved in the DNA recombination event. An IH promoter lies immediately upstream of each S region. IH promoters are activated upon binding of transcription factors induced by CD40 signaling, TLR signaling, occupancy of receptors by cytokines (such as IL-4, IFN-γ, or TGF-β), or BCR crosslinking by antigen.
The IH promoters that lie upstream of the S regions that will be involved in the CSR event are activated to induce germline IH -CH transcripts, which are then spliced at the I region to join with the corresponding C region (FIGURE 16.23).
FIGURE 1. When transcription separates the strands of DNA, one strand forms a single-stranded loop if 5′-AGCT-3′ motifs in the same strand are juxtaposed.
S regions vary in length, as defined by the limits of the sites involved in recombination, from 1 to 10 kb. They contain clusters of repeating units that vary from 20 to 80 nucleotides in length, with the major component being 5′-AGCT-3′ repeats. The CSR process continues with the introduction of DSBs in S regions followed by rejoining of the cleaved ends. The DSBs do not occur at obligatory sites within S regions, because different B cells expressing the same Ig class have broken the upstream and downstream S regions at different points, yielding different recombined S-S sequences.
Ku70/Ku80 and DNA-PKcs, which are required for the joining phase of V(D)J recombination and for NHEJ in general, are also required for CSR, indicating that the CSR joining reaction uses the NHEJ pathway. CSR can occur, though, albeit at a lower efficiency, in the absence of XRCC4 or DNA ligase IV, suggesting that an alternative end joining (A-EJ) pathway can be used in the ligation of S region DSB ends.
A-EJ in CSR entails inclusion of nucleotide microhomologies at S–S junctions, a signature of microhomology-mediated end-joining (MMEJ). The microhomology-mediated A-EJ in CSR is mediated by HR factor Rad52, a DNA-binding element that promotes annealing of complementary DSB single-strand ends. Rad52 competes with Ku70/Ku80 for binding to S region DSB free ends.
There, it facilitates a DSB synaptic process which favors intra-S region recombination. It also mediates, particularly in the absence of a functional NHEJ pathway, inter-S–S region recombinations. The key insight into the mechanism of CSR has been the discovery of the requirement for the enzyme activation-induced (cytidine) deaminase (AID). In the absence of AID, CSR aborts before the DNA nicking or breaking stage. SHM is also abrogated, revealing
an important connection between these two processes, which are central to the maturation of the antibody response and the generation of high-affinity antibodies (see the section in this chapter titled SHM Is Mediated by AID, Ung, Elements of the Mismatch DNA Repair Machinery, and Translesion DNA Synthesis Polymerases).
AID is expressed late in the natural history of a B lymphocyte, after the B cell encounters the antigen and differentiates in germinal centers of peripheral lymphoid organs, restricting the processes of CSR and SHM to this stage. AID deaminates deoxycytidines in DNA and possesses structural similarities to the members of APOBEC proteins that act on RNA to deaminate a deoxycytidine to a deoxyuridine . The expression and activity of AID are tightly regulated at multiple levels. Transcription of the AID gene (Aicda) is modulated by multiple transcription factors, such as the homeodomain protein HoxC4 and NF-κB.
HoxC4 expression is upregulated by estrogen receptors, resulting in upregulation of AID and potentiation of CSR and SHM in antibody and autoantibody responses.
Ung is another enzyme that is required for both CSR and SHM.
Ung, a uracil-DNA glycosylase, deglycosylates the deoxyuridines generated by the AID-mediated deamination of deoxycytidines to give rise to abasic sites. B cells that are deficient in Ung have a 10-fold reduction in CSR, suggesting that the sequential intervention of AID and Ung creates abasic sites that are critical for the generation of DSBs. Different events follow in the CSR and SHM processes.
AID more efficiently deaminates deoxycytidine in DNA that is being transcribed and that, therefore, exists as a functionally singlestrand DNA, such as in germline IH -CH transcription, in which the S region nontemplate strand of DNA is displaced when the bottom strand is used as a template for RNA synthesis (FIGURE 2). Although this has been proposed as an operational model for DNA deamination by AID, it would not explain how AID deaminates both DNA strands, which it does. The abasic site emerging after sequential AID-mediated deamination of deoxycytidine and Ungmediated deglycosylation of deoxyuridine is attacked by an apyridinic/apurinic endonuclease (APE) or MRE11/RAD50, which creates a nick in the DNA strands. Generation of nicks in a nearby location on opposite DNA strands would give rise to DSBs in S regions. The DSB free ends in upstream and downstream S regions are joined by NHEJ (see the section Nonhomologous End- Joining Also Repairs Double-Strand Breaks in the Repair Systems chapter). Aberrant repair of the DSBs would lead to chromosomal translocations. How the CSR machinery specifically targets S regions, and what determines the targeting of the upstream and downstream S regions recruited into the recombination process, is just starting to be understood. 14-3-3 adaptor proteins are involved in recruiting/stabilizing AID to S regions by targeting 5′-AGCT-3′ repeats in S regions. 5′-AGCT-3′ repeats account for
more than 40% of the “core” of S regions and constitute the primary sites of DSBs. Accessibility of S regions by 14-3-3, AID, and other elements of the CSR machinery is dependent on germline IH -CH transcription and chromatin modifications, including histone posttranslational modifications (PTMs). In certain pathological conditions, such as cancer and autoimmunity, AID offtargeting (i.e., targeting of DNA by AID outside the Ig loci) occurs in the genome at large, leading to widespread DNA lesions, such as DSBs, aberrant chromosomal recombinations, and accumulation of mutations in genes that are not physiologically targets of SHM.
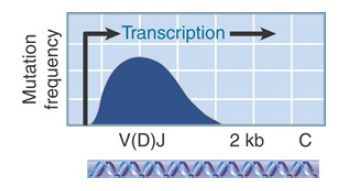
FIGURE 2. Somatic mutation occurs in the region surrounding the V segment and extends over the recombined V(D)J segment.



|
|
|
|
التوتر والسرطان.. علماء يحذرون من "صلة خطيرة"
|
|
|
|
|
|
|
مرآة السيارة: مدى دقة عكسها للصورة الصحيحة
|
|
|
|
|
|
|
نحو شراكة وطنية متكاملة.. الأمين العام للعتبة الحسينية يبحث مع وكيل وزارة الخارجية آفاق التعاون المؤسسي
|
|
|