























النبات

مواضيع عامة في علم النبات
الجذور - السيقان - الأوراق
النباتات الوعائية واللاوعائية
البذور (مغطاة البذور - عاريات البذور)
الطحالب
النباتات الطبية
الحيوان
مواضيع عامة في علم الحيوان
علم التشريح
التنوع الإحيائي
البايلوجيا الخلوية
الأحياء المجهرية
البكتيريا
الفطريات
الطفيليات
الفايروسات
علم الأمراض
الاورام
الامراض الوراثية
الامراض المناعية
الامراض المدارية
اضطرابات الدورة الدموية
مواضيع عامة في علم الامراض
الحشرات
التقانة الإحيائية
مواضيع عامة في التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحيوية والميكروبات
الفعاليات الحيوية
وراثة الاحياء المجهرية
تصنيف الاحياء المجهرية
الاحياء المجهرية في الطبيعة
أيض الاجهاد
التقنية الحيوية والبيئة
التقنية الحيوية والطب
التقنية الحيوية والزراعة
التقنية الحيوية والصناعة
التقنية الحيوية والطاقة
البحار والطحالب الصغيرة
عزل البروتين
هندسة الجينات
التقنية الحياتية النانوية
مفاهيم التقنية الحيوية النانوية
التراكيب النانوية والمجاهر المستخدمة في رؤيتها
تصنيع وتخليق المواد النانوية
تطبيقات التقنية النانوية والحيوية النانوية
الرقائق والمتحسسات الحيوية
المصفوفات المجهرية وحاسوب الدنا
اللقاحات
البيئة والتلوث
علم الأجنة
اعضاء التكاثر وتشكل الاعراس
الاخصاب
التشطر
العصيبة وتشكل الجسيدات
تشكل اللواحق الجنينية
تكون المعيدة وظهور الطبقات الجنينية
مقدمة لعلم الاجنة
الأحياء الجزيئي
مواضيع عامة في الاحياء الجزيئي
علم وظائف الأعضاء
الغدد
مواضيع عامة في الغدد
الغدد الصم و هرموناتها
الجسم تحت السريري
الغدة النخامية
الغدة الكظرية
الغدة التناسلية
الغدة الدرقية والجار الدرقية
الغدة البنكرياسية
الغدة الصنوبرية
مواضيع عامة في علم وظائف الاعضاء
الخلية الحيوانية
الجهاز العصبي
أعضاء الحس
الجهاز العضلي
السوائل الجسمية
الجهاز الدوري والليمف
الجهاز التنفسي
الجهاز الهضمي
الجهاز البولي
المضادات الميكروبية
مواضيع عامة في المضادات الميكروبية
مضادات البكتيريا
مضادات الفطريات
مضادات الطفيليات
مضادات الفايروسات
علم الخلية
الوراثة
الأحياء العامة
المناعة
التحليلات المرضية
الكيمياء الحيوية
مواضيع متنوعة أخرى
الانزيمات

Dihybrid Crosses
 المؤلف:
AN INTRODUCTION TO PLANT BIOLOGY-1998
المؤلف:
AN INTRODUCTION TO PLANT BIOLOGY-1998
 المصدر:
JAMES D. MAUSETH
المصدر:
JAMES D. MAUSETH
 الجزء والصفحة:
الجزء والصفحة:
 19-10-2016
19-10-2016
 2905
2905




+
-
20
Dihybrid Crosses
A dihybrid cross is one in which two genes are studied and analyzed simultaneously, rather than just one, as in a monohybrid cross. Every cross involves all the genes in the organism, but the terms "monohybrid" and "dihybrid" refer only to the number being analyzed.
When two genes are studied, the results of the crosses depend upon the positions of the genes on the chromosomes. If they are on different chromosomes, the alleles for one gene move independently of the alleles for the other gene, but when two genes are close together on the same chromosome, the alleles for one gene are chemically bound to the allele for the other gene and move together. The situation in which the genes are on separate chromosomes is easier to understand and is explained first.
GENES ON SEPARATE CHROMOSOMES: INDEPENDENT ASSORTMENT
Consider a plant heterozygous for two traits, for instance seed coat texture and color, with a smooth seed coat (S) showing complete dominance over a wrinkled seed coat (s), and a yellow seed coat (Y) being dominant over a green seed coat (y). The plant's genotype is SsYy and its phenotype is smooth yellow seeds. We know that if we consider only the gene for color, the plant produces two types of gametes in approximately equal numbers, some carrying Y and some carrying y Also, if we consider only texture, half carry S and half s. How do the alleles of the two genes relate to each other? As in monohybrid crosses, knowing the types of gametes that can be formed is the key to understanding the patterns of inheritance.
If the two genes are on separate chromosomes, the alleles of one gene move independently of the alleles of the other gene during meiosis I; this is called independent assortment. All chromosomes align on the metaphase plate during metaphase I; the chromosomes have duplicated and each has two copies of each allele (Fig. 1a). The homologous chromosomes have also paired, so there are two Y alleles, two y alleles, two S alleles, and two s alleles at the metaphase plate. During anaphase I, homologous chromosomes separate from each other, and both Y alleles move to one pole because they are on the two chromatids of one chromosome, still held together by its centromere. Both y allele, located on the two chromatids of the homologous chromosome, move to the other pole There is no way to predict which pole will receive which type of allele. Similarly, the S alleles separate from the two s alleles and move randomly to the poles. In some cells, the pole that receives the Y alleles at telophase I also receives the S alleles, but in other cells the Yand s alleles end up together. During meiosis II, the two chromatids of each chromosome separate from each other, resulting in four types of haploid cells in equal numbers: SY, sY Sy and sy. Any single microspore or megaspore mother cell produces only two types of haploid cell; the set SY and sy or the set sY and Sy. All four types are produced by a single plant because some mother cells produce one set and some produce the other set.

FIGURE 1: (a) During anaphase I, chromosomes move independently of each other, so one pole receives S and the other receives s; likewise, one gets Yand the other y. But in about half the cells both S and Y move to the same spindle pole by chance, whereas in the other dividing cells, one spindle pole receives S and y, again by chance. (b) A Punnett square for a dihybrid cross is set up just like ere for a monohybnd cross; establish the types and relative abundance of gametes, then fill in the squares. The table looks a little formidable, but it really consists of two 3 : 1 ratios intermingled.
Once the possible types of gametes are known, the Punnett square can be set up, as in Figure 16.18b. Any single fertilization results from the syngamy of one sperm cell and one egg cell and produces only one of the 16 possible zygote genotypes shown in Figure 16.18b. All 16 types of zygote occur only if we study many fertilizations; to have the ratios come out accurately, we have to analyze hundreds of progeny. In plants, it is usually easy to obtain large numbers of fertilizations because pollen and ovules are produced in large amounts. Once pollinated, most plants produce dozens or even thousands of seeds, enough progeny that all 16 zygote genotypes occur in about the expected ratios. But in large animals such as mammals, reproduction may be infrequent and only one or two offspring are produced each year; a great deal of work is necessary to get enough progeny to verify the results of a dihybrid cross.
In a dihybrid cross involving independent assortment of two heterozygous genes, each gene showing complete dominance, a characteristic phenotype ratio occurs, just as is true of the 3; 1 ratio in a monohybnd cross. The ratio is 9:3:3:1, with 9/16 of the plants having the dominant phenotype for both traits (in our example smooth yellow seed coats), 3/16 with the dominant phenotype of the first trait and the recessive phenotype of the second (smooth, green), 3/16 with the first trait recessive and the second dominant (wrinkled, yellow), and 1/16 in which the plants have the recessive phenotype of both traits (wrinkled, green) (Table 16.3). The 9:3:3: 1 ratio results only if all four types of gametes are produced in equal numbers and have equal opportunity to participate in reproduction. The alleles Y and y must be independent of S and s during meiosis I; this automatically happens if they are on different chromosomes.

Notice that if only one trait is considered, it behaves as in a monohybrid cross: Plants with smooth seeds outnumber those with wrinkled seeds by 3:1 and those with yellow seeds are three times more abundant than those with green seeds. Similarly, the monohybrid genotype ratios are also present— 1 SS: 2 Ss: 1 ss and 1 YY: 2 Yy. 1 yy. Considering two genes simultaneously does not affect their inheritance at all.
CROSSING-OVER
Independent assortment can also occur if two genes are located far apart on the same chromosome such that crossing-over occurs between them during prophase I, after homologous chromosomes have paired and a synaptonemal complex is formed . Because no preferential sites for crossing-over seem to exist, the farther apart two genes are, the greater the possibility that crossing-over will occur between them. Most plant chromosomes are so long that crossing-over occurs several times within each chromosome during each prophase I. Consequently, the two ends act like separate entities; if the gene for seed coat color were at one end of the chromosome and the gene for seed coat texture were at the other, they would still undergo independent assortment. However, if the two genes are close together on a chromosome, crossing-over may not occur and the two alleles may move together during meiosis I, as described below.
GENES ON THE SAME CHROMOSOME: LINKAGE
If two genes occur close together on a chromosome, they usually do not undergo independent assortment during meiosis I; instead, the two genes are said to be linked. Consider a plant heterozygous for the two traits of the seed coat again, but now imagine the genes occurring close together on one chromosome. During meiosis, haploid cells that are either S or s, Y or y are formed, but a new complexity arises. Because the genes are linked, several types of heterozygote are possible: an SsYy individual may have the alleles S and Y on one chromosome and the alleles s and y on the homologous chromosome. But an SsYy individual may have s and Y linked together and S and y linked (Fig. 2a). We must consider the types of gametes that can he formed. II crossing-over is ignored, the first individual produces haploid cells with the genotypes SY and sy only, whereas the second plant would produce sY and Sy gametes only. The result of selfing is not a 9 : 3 : 3 :1 ratio, but something drastically different.
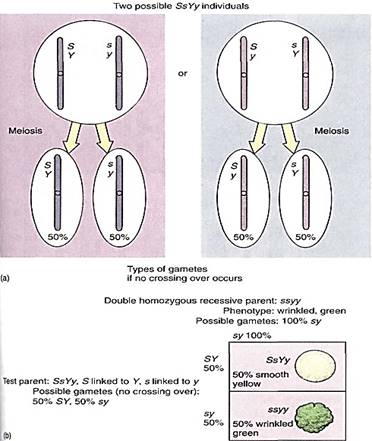
FIGURE 2 (a) Two types of SsYy individuals are possible, but in most instances their phenotypes are identical. Only genetic tests can distinguish which is which, based on the unique gametes produced by each. (b) A test cross, with the plant on the left in (a) being the test parent; try setting up the Punnett square for a test cross with the plant on the right in (a). A test cross of two closely linked genes produces results very different from those of a test cross involving non-linked genes that assort independently. What would the Punnett square be like if S and Y, s and y were not linked?
The most instructive cross is a test cross using a double homozygous recessive pareent: ssyy. If we continue to exclude crossing-over (imagine that the two genes are extremely close together), the gametes from the first type of heterozygote will be SY and sy and the gametes from the homozygous recessive parent will be all sy. Two types of F1 will be produced, as shown in Figure 16.19b, smooth yellow and wrinkled green, and they occur in a ratio of 1 to 1. The two phenotypes are like those of the parents.
If the genes are not extremely close together, crossing-over may occur; lor example, in 10% of the cells undergoing meiosis, a crossing-over might happen, so the plant would produce four types of gametes, but not in equal numbers. Of the gametes for the first type of heterozygote, 47.5% would be SY, 47.5% sy, 2.5% sY, and 2.5% Sy (Fig. 3a). The last two are recombinant chromosomes formed from a crossing-over of the homologous chromosomes and recombination of alleles. The first two are parental type chromosomes. In the majority of the cells, Y is still linked to S and y is linked to s, but in a minority, Y is linked to s and y to S. A Punnett square alone is not sufficient to show the relative proportions of progeny, so the percentages of gamete must be added. A test cross now wild result in 47.5% of the F1 progeny being smooth and yellow, 47.5% wrinkled and green, 2.5% wrinkled yellow, and 2.5% smooth green (Fig. 16.20b). The first two types are like the parents, but the last two are recombinant types produced at the same percentage as crossing-over occurs. We cannot actually "see" crossing-over with a microscope, but we can infer that because 5% of the F1s have a recombinant phenotype, then 5% of the chromatids must have undergone crossing-over.
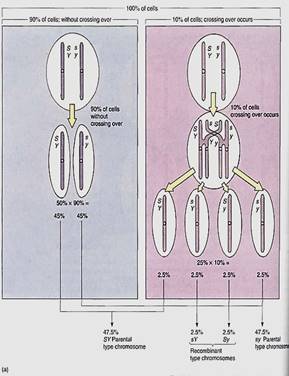
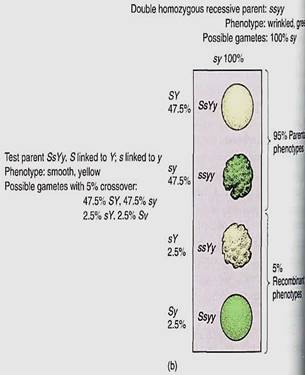
FIGURE 16.20 (a) With linkage, four types of haploid genotypes might be produced, but not in equal numbers. Recombinant types are less abundant than parental types—in this case 5% recombinant chromosomes and 95% parental type. These two genes are 5 map units apart. (b) This test cross, unlike that of Figure 16.19b, gives four types of progeny: two parental types and two recombinant types. The alleles 5 and Y show linkage rather than independent assortment.
The rate of crossing-over is directly proportional to the physical spacing between the genes on a chromosome. Two genes that produce 6% recombinant F1s are closer together than two genes that produce 10% recombinant F1s. By analyzing as many mutant alleles as possible, we can measure "space" between them in recombination percentages, each 1% being called one map unit or one centimorgan; one map unit, on average, is about one million base pairs. By this means, a genetic map can be constructed. If enough genes can be mapped, we may find that A is linked to B which is linked to C; even if A and C are so far apart that they assort independently, by knowing that both are linked to a common gene, B, we know that all three are on the same chromosome. On the other hand, if A, B, and C cannot be shown to be linked to another gene F, that does not mean that F is on a different chromosome; it might be that intervening genes have not yet been mapped (Fig. 4). A set of genes known to be linked is called a linkage group; when all genes are mapped, there will be exactly as many linkage groups as there are chromosomes. But if only a few genes are mapped, there may be more linkage groups than chromosomes.
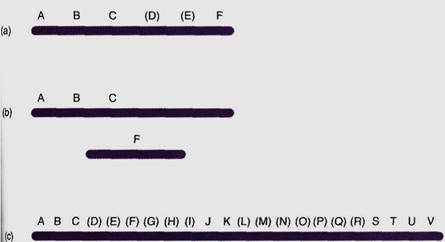
FIGURE 4: (a) A and C might be far enough apart for crossing-over to occur in virtually 100% of the nuclei, so they undergo independent assortment. If each can be shown to be linked to B, they must be on the same chromosome. If A, B, and C are not known to be linked to F, it might be because they are on separate chromosomes (b) or because they are on the same chromosome but intervening genes have not yet been mapped, as shown in (a). (c) Because of incomplete mapping, ABC, JK, and STUV are three linkage groups even though there is only one chromosome.
 الاكثر قراءة في مواضيع عامة في علم النبات
الاكثر قراءة في مواضيع عامة في علم النبات
 اخر الاخبار
اخر الاخبار
اخبار العتبة العباسية المقدسة

الآخبار الصحية


مواضيع ذات صلة
 قسم الشؤون الفكرية يصدر كتاباً يوثق تاريخ السدانة في العتبة العباسية المقدسة
قسم الشؤون الفكرية يصدر كتاباً يوثق تاريخ السدانة في العتبة العباسية المقدسة "المهمة".. إصدار قصصي يوثّق القصص الفائزة في مسابقة فتوى الدفاع المقدسة للقصة القصيرة
"المهمة".. إصدار قصصي يوثّق القصص الفائزة في مسابقة فتوى الدفاع المقدسة للقصة القصيرة (نوافذ).. إصدار أدبي يوثق القصص الفائزة في مسابقة الإمام العسكري (عليه السلام)
(نوافذ).. إصدار أدبي يوثق القصص الفائزة في مسابقة الإمام العسكري (عليه السلام)



















