خطوات هندسة الكائنات الحية وراثياً
قبل ان نخوض ونتعمق في هندسة الكائنات الحية وراثيا يجب التطرق الى بعض المفاهيم الخاصة بتلك التقنيات.
أولا : تصميم الخرائط الوراثية Genetic Maps Design
ويقصد به تحديد الموقع الطبيعي لجين ما او العلاقة الوراثية على كروموسوم ما، والخريطة الخطية للمواقع النسبية للجينات على امتداد الكروموسوم. ويمكن التعرف على المسافات الجينية من خلال تحليل الارتباط، والتي تحدد التكرار والتي عندها تعتبر المواقع الجينية منفصلة اثناء اعادة التوليف الكروموسومي. ويمكن تشبيه تصميم الخرائط الوراثية بالعرض البياني المركز للمسافات النسبية ولكن معبرا عنها بالاتحادات الجديدة بين جينات المجموعات الارتباطية الواحدة والمحمولة على كروموسوم واحد والمقصود برسم الخرائط الوراثية هو تحديد المواقع النسبية لمقاطع المادة الوراثية DNA fragments المختلفة في المحتوى الوراثي لكائن وتحديد مدى ارتباط هذه المقاطع بالصفات الوراثية سواء الكمية التي تعتمد في توارثها على العديد من الجينات مثل كمية المحصول او النوعية التي تعتمد في توارثها على جين واحد او عدد قليل من الجينات.
وتسمى هذه المقاطع من المادة الوراثية بالجينات وتلعب الخرائط الوراثية دور بارز في برامج التربية والتحسين الوراثي فهي تعتبر المرشد الذي عن طريقه يمكن ان يبدا المربي برنامجه بخطي ثابتة واثقة امنة حتى يصل الى الهدف المنشود في اقصر وقت ممكن. فمثلاً اذا استطعنا ان نحدد مقطع او مقاطع معينة من المادة الوراثية DNA يرتبط ظهوره بوجود صفة اقتصادية هامة مثل المقاومة لمرض معين او زيادة كمية المحصول... الخ. وعلى أيه حالة، فان طريق اجراء اختبارات على مستوى DNA باستخدام تقنيات البيولوجيا الجزيئية يمكن انتخاب النباتات الحاملة لهذه المقاطع والتي ترشد المربي الى وجود الصفة المرغوب فيها مباشرة وبدقة مما يمكنه من الوصول الى الهدف المنشود من برنامج التربية من خلال جيلين او ثلاثة بدلا من 1 الى 15 جيلا باتباع الطرق التقليدية.
1- الارتباط الوراثي او الخرائط الهجينة Genetic Linkage or Hybrid Maps
تبنى الفكرة في الارتباط الوراثي (الخرائط الارتباطية) على انه عند التهجين بين نباتين او أي كائنين فان نتائج التلقيحات بين ازواج الاليلات الجينية تكون 50% تراكيب ابوية واقل من 50% تراكيب ذات اتحادات جديدة فاذا ما ظهرت النتائج لكثير من 50% تركيب ابوية واقل من 50% اتحادات جديدة دل ذلك على وجود ارتباط لتلك الصفات او بمعنى اخر وجود الجينات المسئولة عن تلك الصفات على كروموسوم واحد. وحيث ان الاتحادات الجديدة تحدث عند حدوث العبور بين الموقعين التي تقع فيهم الجينات فان احتمال حدوث العبور يتوقف على المسافة التي تفصل بين الجينات على الكروموسوم.
2- تهجين الأحماض النووية وخرائط التماثل Nucleic Acid Hybridization and Homology Map
هناك تقنيات حديثة تسمى تهجين الاحماض النووية Nucleic acid hybridization and homologies وهي طريقة يستدل بها على ترتيب او تسلسل معين من القواعد النيتروجينية او على جين معين فعند تسخين معلق من DNA مزدوج السلسة والمحتوى على مجموعة من الجينات غير المعروفة الوظيفة فان الروابط الهيدروجينية بين ازواج القواعد النيتروجينية تنكسر فتنفصل كلتا السلسلتين في كل جين وعند تبريد المعلق المحتوى DNA فإن السلسلتين تعاودا الارتباط لان كل منهما يميلا لتكملة بعضهما البعض تماما، فاذا تم خلط DNA من خلية ما مع DNA مصنع او الحامض النووي mRNA مصنع حيث يعزل البروتين المسبب لظاهرة فسيولوجية معينة ويتم دراسة تسلسل وترتيب الاحماض الامينية به وتعطى تلك البيانات الى الكمبيوتر لاستنباط وتوقع تسلسل الجين المسئول عن هذا البروتين ثم يقوم جهاز PCR بتكوين شظايا الحامض النووي DNA او RNA المتوقع، لكن نظرا لوجود عدة احتمالات لتتابع النيوكليوتيدات داخل الجين محل الدراسة ونظرا لان الشفرة الوراثية للأحماض الامينية ليست شفرة واحدة فللحمض الاميني اكثر من شفرة واحدة فاذا ما خلط DNA المصنع وحدث الارتباط بين الجين المطلوب البحث عنه مع احدى جزيئات DNA المصنع سمى الناتج Homologous DNA وتسمى تلك التقنية باسم DNA Hybridization ويمكن جعل احد الاحماض النووية الداخلة في تركيب DNA المستخدم في التعرف على الجين مشع Radioactive بذلك يمكن اصطياده ومن طبق الزراعة Petri dish الذي يتم تحضينه تحت التبريد لمدة 24 ساعة وعليه لوحة من فيلم حساس ليظهر عليها تأثير القواعد المشعة لنتعرف على الجينات التي حدث لها تهجين فيتم عزلها وتنقيتها واكثارها ثم استخدامها في هندسة نبات آخر.
3- الخرائط الجزيئية Molecular Maps
تعد خاصية اعادة الاتحاد ذات فائدة كبيرة في البيولوجيا الجزيئية حيث تستخدم في قياس طول الكروموسوم ومعرفة عدد النيوكلوتيدات حيث انه تحت الظروف القياسية فان الجينوم الاكبر حجما سيأخذ وقتا اطول في اعادة الاتحاد عن الجينوم الاصغر، ومن معرفة الزمن يمكن تحديد طول الكروموسوم وعدد نيكليوتيداته، كما يمكن رسم خريطة لجزئ DNA اعتمادا على حقيقة ان المناطق المحتوية على T, A تنفصل بمعدل اسرع نتيجة احتوائه على زوجين من الروابط الهيدروجينية ويمكن التعرف على هذه المناطق تحت الميكروسكوب الالكتروني على شكل عروات Loops أو فقاعات Bubbles كما يمكن قياس المسافات بين العروات ايضا وبين نهاية جزيئي DNA.
4- استخدام الميكروسكوب الالكتروني في رسم الخرائط الكروموسومية
عند الحصول على طفرة ما في النبات او الحيوان ونريد التعرف على مكانها لتساعدنا على رسم الخرائط الوراثية، يتم عزل DNA من كل من النبات الطافر والنبات السليم باستخدام حمام مائي على درجة 100° م وباستخدام مادة قلوية لترفع PH الى 11.5 فيتم هدم DNA الى سلسلتين منفصلتين وعند خلط DNA من كل من النباتين الطافر والسليم يتم اعادة اتحاد السلسلتين ولكن بصورة خليطة ويحدث انبعاج لاحد الخيطين single stranded loops لعدم تمكن الازدواج في الجين الطافر بين احد السلسلتين السليمة مع السلسلة الطافرة والتي يمكن تحديد مكانها على أي من الكروموسومات وطولها بواسطة الميكروسكوب الالكتروني.
5- خرائط الانتشار الانزيمي المقيد Restriction Fragment Length Polymorphism
تستخدم فيها انزيمات القطع المحددة او انزيمات الاندونيوكليز التي تقطع جزئ DNA في اماكن محددة عند تتابعات معينة من النيوكلوتيدات حيث قام العالمان Smith & Nathans بتوصيف تلك الانزيمات وقد حازا على جائزة نوبل عام 1978 لاكتشافهما انزيمات الاندونيوكليز المقيدة وباستخدام تلك الانزيمات يمكن تقطيع DNA الى قطع ويمكن تحديد حجم كل قطعة باستخدام نظام التفريد الكهربي باستخدام جيل البولي اكريميليد او الاجاروز polyacrylamide او Agarose نظرا لان كل وحدة كروماتيدية سوف تحمل شنة سالبة ناتجة عن مجموعة الفوسفات وعليه فان معدل الهجرة لقطع DNA خلال عملية التفريد الكهربي يعطي مقياس دقيق لأطوالها حيث يتناسب معدل الهجرة عكسيا ونسبيا مع طولها، وتصبغ قطع DNA بصبغة الاثديوم بروميد Ethidium bromide حيث تربط الصبغة DNA وعند تعرضها للأشعة فوق البنفسجية يظهر DNA كوميض فلورسنتي فيسهل تعيينها وتصويرها وسوف نتناول ذلك بشيء من التفصيل لاحقا.
ثانيا : دراسة تتابع النيوكلوتيدات داخل الجين Ultimate Structure Maps
لمعرفة التركيب المتناهي الدقة للخرائط فان ذلك يتم بمعرفة تتابع النيوكلوتيدات ومعرفة التسلسل النووي داخل الجين حتى يمكن اختيار انزيمات القطع المحدد الواجب استخدامها للحصول على الجين المطلوب وهو ما يعرف باسم الخرائط الجينية فائقة الدقة Ultimate Fine Structure Maps، وقد اكتشف Sanger وزملاءه 1976 طريقة انزيمية ومنهيات لسلسلة DNA لإنتاج قطع من DNA تكون نهايتها عبارة عن نيوكليوتيدات خاصة تحتوي على سكر رايبوزي من نوع خاص هو Dideoxyribolose-2.3 حيث تشتمل ذرتي الكربون الثانية والثالثة على ذرة هيدروجين ناقصة لذرتي الاوكسجين وبدلا من مجموعة OH على ذرة الكربون الثالثة والضرورية جدا ليستطيع انزيم DNA polymerase من العمل على اتحاد النيوكلوتيدات وتكون رابطة الاستر بين مجموعة الهيدروكسيل بالسكر ومجموعة حمض الفوسفوريك في النيوكليوتيدة التالية لتتكون سلسلة DNA الفردية قبل اتحادها مع مثيلتها لتكوين سلسلتين الحامض النووي DNA المزدوج الحلزوني. فاذا عزل جين ما فأريد معرفة تركيبه الدقيق وترتيب نيكليوتيداته، يتم وضع اجزاء DNA لإجراء اربعة تفاعلات متوازية لبناء خيط مكمل له بواسطة انزيمات الاندونيوكليز والتي تقوم بفك حلزون قطع DNA (القالب) ونسخ صورة مراه منه باستخدام اربعة انواع من النيوكلوتيدات؛ ثلاثة منها نيوكليوتيدات عادية والرابعة نيوكليتيدة من نوع 2,3-Dideoxyribolose كمنهيات للتفاعل فيستعمل في التفاعل الاول in vito نيوكليتيدة ddATP وفي التفاعل الثاني نيوكليتيدة ddGTP وفي التفاعل الثالث نيوكليتيدة ddCTP وفي التفاعل الرابع نيوكليتيدة ddTTP وهي ذات نشاط اشعاعي حيث تحتوي على الفوسفور 32 المشع فينتج عن التفاعلات قطع من DNA لكنها متقطعة دائما عند النهايات المستخدمة وعند فصلها بالتفريد الكهربي باستخدام جيل البولي اكريميليد polyacrylamide يمكن تعيين موقعهم في الجيل بواسطة الاشعاع، وبذا سوف ينتج سلما يشير الى تتابعات النيوكلوتيدات وسوف تذهب اقصر القطع الى اطول مسافة او اقرب جهة للانود (الالكترود الموجب) وستكون كل حزمة تالية محتوية على سلاسل اطول وهكذا قراءة السلم في جيل polyacrylamide المستعمل في الفصل.
ثالثا : معالجة الجين المعزول لكي يعبر وراثيا عن نفسه Gene Expression
لكي يتم تعبير الجين وراثيا أي نسخ الجين لنفسه وتكوين صورة على شكل الحامض النووي mRNA ليتم ترجمتها على الرايبوسومات لتكوين البروتين اللازم لإظهار صفة نباتية مرغوبة (شكل ظهري Phenotype) يجب ان يتكون هذا الجين من ثلاثة مناطق :
- المنطقة الاولى تسمى تسلسل المحفز Promoter sequence وهي تساعد في تحديد توقيت عمل الجين وموقع تعبير الجين فيه بمثابة شفرة للجين نفسه وتحدد مكان بدء نسخ الحامض النووي RNA الرسول (ابدأ من هنا) (شكل 1).
- المنطقة الثانية هي منطقة التشفير وهي تحمل معلومات تحديد طبيعة البروتين الذي يشفره الجين التركيبي Structured gene.
- المنطقة الثالثة والتي يطلق عليها منطقة الادينين المتعددة Ploy adenylation (Poly-A ) وهي المسئولة عن انهاء عمل نسخة الحامض النووي tRNA الرسول للحامض النووي RNA transcript Messenger على الوجه الصحيح وكأنها تقول للجين انهى عملية النسخ هنا.
ولحسن الحظ ان امام المتخصص في الهندسة الوراثية حرية واسعة في مزج هذه المناطق والموائمة بينما وتجميعها من جينات مختلفة لتنتج ما يسمى بالجينات الكيميرية او الخليطية Chimeric genes وبذلك امكن للمتخصصين في الهندسة الوراثية اختيار محفزات متباينة فامكنهم توجيه تعبير الجين الى اعضاء بذاتها مثل الاوراق او البذور او الجذور او الدرنات بل الى انماط بذاتها من الخلايا داخل النسيج الواحد. وقد يصمم الجين الكيميري او الخليطة من جينات كائنات مختلفة فالمحفز من فيروس نباتي ومنطقة التشفير من بكتيريا E. coli موقع تعدد الادننة poly A من Agrobacterium ثم يتم الايلاج في خلية نباتية تقوم بنسخ الحامض النووي mRNA لتترجمه الرايبوسومات ribosomes لتنتج البروتينات .
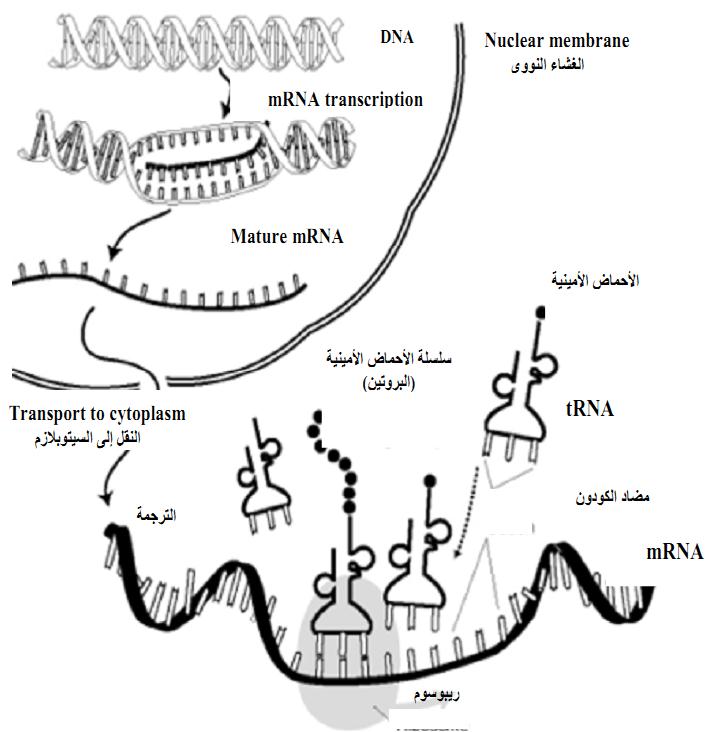
شكل 1 : التعبير الجيني Gene Expression
رابعا : مرحلة تطعيم الجين واكثاره Gene Cloning
تأتي مرحلة تطعيم الجين الذي تم تركيبة على بلازميد خلية بكتيرية والبلازميدات هي تراكيب وراثية غير كروموسوميه للبكتيريا وهي عبارة عن جزيئات من DNA تتضاعف مستقلة عن الكروموسوم في النواة غير الحقيقية وتحتوي تلك البلازميدات على موروثات تمكنها من الانتقال من الخلية المانحة Donor cell الى خلية اخرى لذلك تسمى تلك البلازميدات بالبلازميدات المعدية او بلازميدات الاتصال . وقد تتصل بعض البلازميدات بكروموسوم الخلية عن طريق عملية اتصال مزدوج (عبور وراثي ) وعند اتصال البلازميد مع كروموسوم الخلية فانة لا يتضاعف او ينسخ مستقلا بذاته بل يصبح تضاعفه مرتبطا بتضاعف الكروموسوم ثم بعد التضاعف تعيد استقلاليتها عن الكروموسوم وهي نفس خاصيه الفيروسات المعتدلة Temperate Viruses والتي تستخدم في هندسة الكائنات الاخرى وراثيا عبر النهايات اللاصقة لأنزيم قطع واحد حيث يتم القطع بنفس المكان من التتابع فيؤدي ذلك الى تكوين نهايات لاصقة يمكن بواسطتها لحام قطعة من DNA المعزول الى جينوم الكائن المهندس وراثيا .
ويمكن التخلص من البلازميدات بعد قيامها بدورها كناقل Vector دون قتل الخلايا الحاوية للبلازميدات وذلك بعملية المعالجة curing عن طريق تثبيط انقسام البلازميد اثناء انقسام الخلايا النباتية مثلا في معلق الخلايا فيتم تخفيف البلازميدات في المستعمرة وذلك باستخدام ملح بروميد الايثديم Ethidium bromide ولقد امكن استغلال البلازميدات كناقل لإدخال الجينات في جينوم كائن اخر حيث ان البلازميد تخترق النواة عند انقسامها وتقوم بفرد حلقتها ولصقها بإحدى كروموسومات النواة فعند تضاعف الكروموسوم اثناء الانقسام يتم عمل نسخة اضافية من البلازميد الذي سرعان ما ينفصل عن الكروموسوم ويخرج من النواة الى السيتوبلازم الجديد مرة اخرى ، ويتم اختيار البلازميد الذي يحتوي على علامة كأن يكون محتوى على جين لمقاومة للاستربتوميسين او ينقل للبلازميد جين مقاوم للمضاد الحيوي كاناميسين ثم يتم فتح حلقة البلازميد باستخدام احدى انزيمات القطع المحددة وينقل اليها الجين الجديد المرغوب اكثاره وتطعيمه عن طريق انزيم اللصق او اللحام الليجيز ، ثم يتم التحام الجين الجديد بحلقة البلازميد ثم يتم ادخال البلازميد المطعوم الى داخل البكتيريا E.coli او Agrobacterium tumefaciens التي تتكاثر بسرعة هائلة فيتم مضاعفه عدد البلازميدات المحتوية على الجين الجديد ولتميز الخلايا البكتيرية المحتوية على بلازميد المطلوب يتم معاملتها بأحد المضادات الحيوية استربتوميسين او الكاناميسين فخلايا البكتيريا التي تقاوم تكون هي المحتوية على الجين المطلوب .
خامسا: نقل الجين الى الجينوم Transformation
هناك طرق عديدة لنقل الجين المعزول وادخالها في جينوم الكائن المرغوب هندسته نوجزها في التالي :
ا- النقل بواسطة بكتيريا Agrobacterium tumefaciens
اول نظام لهندسة النباتات وراثيا وهو الاوسع استخداما هو نقل الجين المرغوب الى النبات باستخدام قدرة البكتيريا Agrobacterium tumefaciens الممرضة في نقل جزء من DNA الى خلايا النبات وتقوم البكتيريا بنقل جزء من DNA لديها (او الخاص بها) تسمى Transferred DNA(tDNA) بالاندماج بكروموسومات النبات المصاب لتدفعه الى انتاج الهرمونات النباتية لترفع مستواها في تلك الخلايا الى المستوى الذي يؤدي الى سرعة تكاثر الخلايا وتكوين كتل من الخلايا والتي تعرف بالتورد القمي Grown Gall (شكل 2) .

شكل 2 : تكاثر الخلايا وتكوين كتل من خلايا بالتورد القمي
ليصبح هذا التورد مكان صالحا وبيئة ملائمة ومصدر غذائي لتلك البكتيريا فيما يعرف بمرض التورد القمي ولكي تكون تلك البكتيريا فعالة كأداة للنقل الجيني لابد من استئصال جيناتها المسببة للمرض بمعنى نزع سلاحها disarming . ولقد نجح Mary Dell Chilton سنة 1983 واخرون من شركة مونسانتو وجامعة واشنطن من استئصال الجينات الممرضة دون المساس بالية نقل DNA وبالرغم من بساطة الطريقة ودقتها الا ان كثير من المحاصيل من بينها محاصيل الحبوب مثل الارز والقمح والذرة ليست من عوائل الاجروبكتريوم لذلك تم البحث عن نظم بديلة .
ب- دمج الجينات الى خلايا البروتوبلاست Competent Cells Technique
في هذه التقنية يزال جدار الخلايا لان ثقوب الخلية الموجودة بجدار الخلية اصغر من ان تسمح لـ DNA بان تمر بسهولة اما عندما يزال الجدار فلن يعيق نقل DNA سوى الغشاء البلازمي والذي يمكن لمركب عضوي مثل البولي اثيلين جليكول (PEG) من تسهيل اختراق DNA للغشاء البلازمي ومن اكثر العوامل المساعدة شيوعا في اداء هذا العمل كما يمكن دمج DNA في خلايا البروتوبلاست بواسطة الثقب الكهربي Electroporation وفي هذه الطريقة تقوم النبضات كهربائية قصيرة بأحداث ثقوب سريعة الزوال في غشاء الخلية العارية يمكن ان تمر جزئيات DNA من خلالها لكم تلك التقنية اي عزل البروتوبلاست وجد انها تقنية صعبة في كثير من الحبوب وينتج عنها نباتات عقيمة .
ج- طريقة الحقن المجهري Microinjection Technique
تتم هذه الطريقة باستخدام ابر خاصة لحقن المادة الوراثية داخل نواة الخلية تحت مايكروسكوب خاص يسمى Micro manipulator واستخدمت تلك الوسيلة في نقل DNA ولكن وجد انها تقنية غير عملية لأسباب عدة منها ان طرف الابرة المستخدمة عادة ما ينسد او ينكسر كما ان ادخال DNA للخلايا عملية مجهدة ولا تلائم العمل التجاري ولأيمكن بها ضمان التحام الجين المنقول الى جينوم الخلية (شكل 3) .
شكل 3: الحقن المجهري
د- تقنية المسدس الجيني Gene Gun Technique
وهي طريقة لقذف الخلايا النباتية بالمادة الوراثية المنقولة بعد تغليفها لجسيمات معدنية فلزية ذات اقطار 1-2 ميكرون مثل كريات الذهب . يتم قذف تلك الجسيمات بسرعة عالية باستخدام المسدس الجيني لتخترق طلاقاته جدار الخلايا وتنقل الجين المرغوب (شكل 4) .

شكل 4: تقنية قاذفات الجسيمات الدناوية DNA
ونظرا لان الثقوب التي يحدثها القذف السريع صغيرة للغاية فهذه الثقوب تكون مؤقتة ولا تعرض سلامة الخلايا للخطر ويتكون المسدس الجيني من قاذف خرطوشي عيار 0.22 مم كقوة دافعة يحتوي على بارود فقط .
هـ - النقل بالفاج Phage Transportation
سميت ظاهرة انتقال صفة وراثية من خلية بكتيرية الى خلية بكتيرية اخرى بواسطة الفاج باسم النقل بالفاج Transudation حيث تندمج قطعة من كروموسوم بكتيري داخل جسم الفاج وعندما يغزو الفيروس خلية عائل جديد فان الخلية المستقبلة تنقل على كروموسومها القطعة التي في حوزة الفيروس ومن اشهرها Lambda phage (شكل 5) .
فعند مهاجمة هذا الفيروس لبكتيريا القولون فان الخلايا تظل فترة وهي في طور القدرة الكامنة للتحلل Lysogenic حيث تحتوي على نسخة من DNA الفيروس المندمج مع كروموسوم الخلية بيم موقع الجين جلكتوز وموقع الجين البيوتين فاذا ما عرضت الخلايا الى الحث Induction فان الفيروس قد ينفصل عن كروموسوم الخلية من نفس النقطة التي اتصل بها بالكروموسوم بعملية عكس عملية الاندماج ليبدا في مضاعفة نفسه وتكوين الفيريونات الكاملة .
قد تنفجر ولكن في حالات اخرى لا ينفصل الفاج الاولى من نفس نقطة الاتصال بل من اماكن اخرى لذا تحتوي حلقته على DNA الفيروس مضافا الية قطعة من كروموسوم الخلية فاذا احتوى الفيروس على الجين الكلاكتوز سمى الفيروس باسم فاج لامبدا الكلاكتوز حيث ان كمية DNA التي يمكن تعبئتها في الغطاء البروتيني للفاج ثابتة ومحددة فان اضافة موروث الكلاكتوز تعنى بالضرورة استبعاد جزء من مورثات إلفاج لامبدا من ناحية اخرى مما يؤدي الى نقصها لصفات وراثية وهو يفسر عدم القدرة على الحصول على فاج يحتوي على اكثر من موروث او جين واحد .
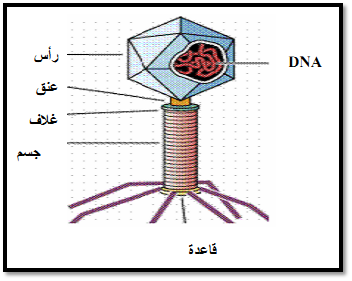
شكل 5: النقل بالفاج لامبدا
وعند استخدام هذه الفاجات لإصابة خلايا البكتيريا ليس لها القدرة على تمثيل الكلاكتوز فتتحول الخلايا الى خلايا لها تلك القدرة على فصل الجينات التي تم انتقالها بالفاج .
سادسا : زراعة الانسجة النباتية Plant Tissue Culture
قد تستخدم اجزاء نباتية Explants بعد معالجتها بالهندسة الوراثية في الاستزراع على بيئات في مزارع اختيارية معقمة وتحفظ معمليا in vitro للحصول على نباتات مهندسة وراثيا بالكامل . كان اجراء الابحاث على الحمض النووي حتى سنة 1970م من اصعب الامور التي كانت تواجه علماء الوراثة والكيمياء . وكانت معظم الابحاث تجرى بشكل غير مباشر على الحمض النووي الريبوزي والبروتين . ولكن تحول الحال بشكل كامل فلصبح علم الوراثة المتعلق بفحص DNA والمعروف بعلم الوراثة الجزيئية من اسهل العلوم واكثرها تطورا . ولقد اصبح من السهل صنع نسخ عديدة من اي جين او مقطع محدد من DNA ، كما امكن معرفة تسلسل الاحماض النووية بسرعة تتعدى المئات في اليوم الواحد . كما استطاع العلماء استكشاف الجينات الموجودة على الكروموسومات كما استطاعوا تغييريها وتعديلها حسب الشكل المراد وليس هذا فحسب بل استطاعوا ان يعيدوا هذه الجينات المعدلة الى الخلية وغرزها في الكروموسوم المراد .
كما امكن انتاج كميات كبيرة من البروتينات كالهورمونات واللقاحات المختلفة والتي كانت تنتج في السابق من جثث الموتى او تستخلص من الحيوانات والتي كانت تحفها بالمخاطر من انتقال العدوى الى الانسان . كما فتحت هذه الثورة العلمية المجال امام الكثيرين من محبي هذا العلم في اختراع واكتشاف طرق جديدة وحديثة في التعامل وحفظ وتغير هذه المادة الحيوية في الانسان والحيوان والنبات . ولقد غير هذا العلم المنطق كالصاروخ الكثير من المفاهيم الطبية والتي دفع كثير من كليات الطب الى تعديل مقرراتها لتزويد طلابها بالمزيد من هذا العلم . ولقد اطلق على عملية نسخ وتعديل وزرع الجينات اسم الهندسة الوراثية Genetic Engineering وهو اسم عام لا يحدد فكرة معينة او تقنية محدده ، ولكنة يعني الكثير من التقنيات وهي متناثرة وموزعة على ا ل كثير من فروع الطب والعلوم .
المصادر:
عبيده ، علي ابراهيم علي و محمود ، احمد عبد الفتاح .اساسيات التقنية الحيوية . كلية الزراعة جامعة الاسكندرية .





















