Tissue-Specific Metabolism: The Division of Labor:- Muscles Use ATP for Mechanical Work
Metabolism in the cells of skeletal muscle—the myocytes—is specialized to generate ATP as the immediate source of energy for contraction. Moreover, skeletal muscle is adapted to do its mechanical work in an intermittent fashion, on demand. Sometimes skeletal muscles must work at their maximum capacity for a short time, as in a 100 m sprint; at other times more prolonged work is required, as in running a marathon or extended physical labor. There are two general classes of muscle tissue, which differ in physiological role and fuel utilization. Slow-twitch muscle, also called red muscle, provides relatively low tension but is highly resistant to fatigue. It produces ATP by the relatively slow but steady process of oxidative phosphorylation. Red muscle is very rich in mitochondria and is served by very dense networks of blood vessels, which bring the oxygen essential to ATP production. It is the cytochromes in mitochondria and the hemoglobin in blood that give the tissue its characteristic red color. Fast-twitch muscle, or white muscle, has fewer mitochondria than red muscle and is less well supplied with blood vessels, but it can develop greater tension, and do so faster. White muscle is quicker to fatigue, because when active, it uses ATP faster than it can replace it. There is a genetic com ponent to the proportion of red and white muscle in any individual; with training, the endurance of fast-twitch muscle can be improved. Skeletal muscle can use free fatty acids, ketone bodies, or glucose as fuel, depending on the degree of muscular activity (Fig. 1). In resting muscle, the primary fuels are free fatty acids from adipose tissue and ketone bodies from the liver. These are oxidized and de graded to yield acetyl-CoA, which enters the citric acid cycle for oxidation to CO2. The ensuing transfer of electrons to O2 provides the energy for ATP synthesis by oxidative phosphorylation. Moderately active muscle uses blood glucose in addition to fatty acids and ketone bodies. The glucose is phosphorylated, then degraded by glycolysis to pyruvate, which is converted to acetyl CoA and oxidized via the citric acid cycle and oxidative phosphorylation. In maximally active fast-twitch muscles, the demand for ATP is so great that the blood flow cannot provide O2 and fuels fast enough to supply sufficient ATP by aerobic respiration alone. Under these conditions, stored muscle glycogen is broken down to lactate by fer mentation (p. 523). Each glucose unit degraded yields three ATP, because phosphorolysis of glycogen produces glucose 6-phosphate, sparing the ATP normally consumed in the hexokinase reaction. Lactic acid fermentation thus responds to an increased need for ATP more quickly than does oxidative phosphorylation, supplementing basal ATP production that results from aerobic oxidation of other fuels via the citric acid cycle and respiratory chain. The use of blood glucose and muscle glycogen as fuels for muscular activity is greatly enhanced by the secretion of epinephrine, which stimulates both the release of glucose from liver glycogen and the breakdown of glycogen in muscle tissue. The relatively small amount of glycogen in skeletal muscle (about 1% of its total weight) limits the amount of glycolytic energy available during all-out exertion. Moreover, the accumulation of lactate and consequent decrease in pH in maximally active muscles reduces their efficiency. Skeletal muscle, however, contains another source of ATP, in the form of phosphocreatine (10 to 30 mM), which can rapidly regenerate ATP from ADP by the creatine kinase reaction:
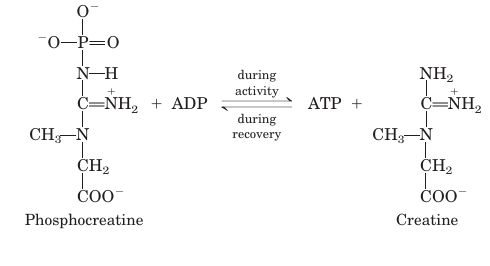
During periods of active contraction and glycolysis, this reaction proceeds predominantly in the direction of ATP synthesis; during recovery from exertion, the same enzyme resynthesizes phosphocreatine from creatine at the expense of ATP.
After a period of intense muscular activity, the in dividual continues breathing heavily for some time, using much of the extra O2 for oxidative phosphorylation in the liver. The ATP produced is used for gluconeogenesis from lactate that has been carried in the blood from the muscles. The glucose thus formed returns to the muscles to replenish their glycogen, completing the Cori cycle (Fig. 2). Heart muscle differs from skeletal muscle in that it is continuously active in a regular rhythm of contraction and relaxation, and it has a completely aerobic metabolism at all times. Mitochondria are much more abundant in heart muscle than in skeletal muscle, making up almost half the volume of the cells (Fig. 3). The heart uses as its fuel mainly free fatty acids, but also some glucose and ketone bodies taken up from the blood; these fuels are oxidized via the citric acid cycle and oxidative phosphorylation to generate ATP. Like skeletal muscle, heart muscle does not store lipids or glycogen in large amounts. It does have small amounts of reserve energy in the form of phosphocreatine, enough for a few seconds of contraction. Because the heart is normally aerobic and obtains its energy from oxidative phosphorylation, the failure of O2 to reach a portion of the heart muscle when the blood vessels are blocked by lipid deposits (atherosclerosis) or blood clots (coronary thrombosis) can cause that region of the heart muscle to die. This is what happens in myocardial infarction, more commonly known as a heart attack.
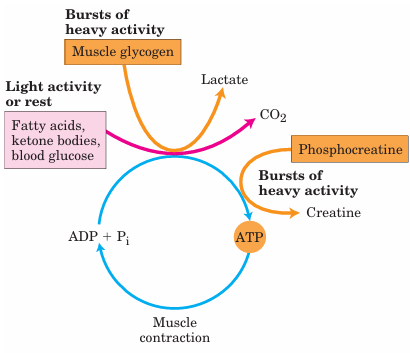
FIGURE 1 Energy sources for muscle contraction. Different fuels are used for ATP synthesis during bursts of heavy activity and during light activity or rest. Phosphocreatine can rapidly supply ATP.
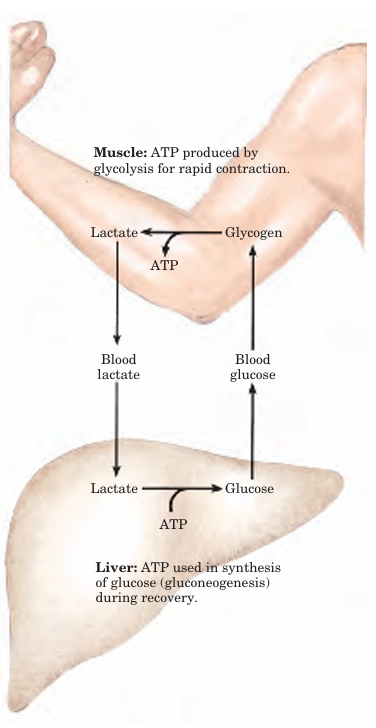
FIGURE 2 Metabolic cooperation between skeletal muscle and the liver. Extremely active muscles use glycogen as energy source, generating lactate via glycolysis. During recovery, some of this lactate is transported to the liver and converted to glucose via gluconeogenesis. This glucose is released to the blood and returned to the muscles to replenish their glycogen stores. The overall pathway (glucose → lactate → glucose) constitutes the Cori cycle.
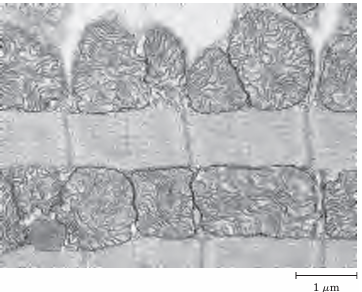
FIGURE 3 Electron micrograph of heart muscle. In the profuse mitochondria of heart tissue, pyruvate, fatty acids, and ketone bodies are oxidized to drive ATP synthesis. This steady aerobic metabolism allows the human heart to pump blood at a rate of nearly 6 L/min, or about 350 L/hr—or 200x106 L over 70 years.





















