Antisense Oligonucleotides
Antisense oligonucleotides are used to hybridize to a specific RNA molecule in vivo and thereby to inhibit its subsequent use. In most cases, the target RNA is a messenger RNA (mRNA), which cannot then be translated into a protein. All that is required is knowledge of the gene's nucleotide sequence, so that an antisense oligonucleotide with the complementary sequence can be synthesized. In this way, the expression of one gene can be blocked, and the consequent observed effects help to elucidate the physiological role of the gene and its product.
Selection of the sites in a RNA molecule at which optimal antisense activity may be induced is complex, dependent on the terminating mechanism and influenced by the chemical class of the oligonucleotide. Each RNA displays a unique pattern of sites of sensitivity. Within the phosphorothioate oligodeoxynucleotide class, studies have shown that antisense activity can vary from undetectable to 100% by shifting an oligonucleotide by just a few bases in the RNA target (1 2, ) . Significant progress has been made in developing general rules that help to define potentially optimal sites in RNA species, but to a large extent this must be determined empirically for each RNA target and every new chemical class of oligonucleotides.
1. Therapeutic Uses
Besides being useful for characterizing the roles of genes, antisense oligonucleotides have the potential for therapeutic use to regulate the expression of certain genes or to block an infection by a pathogenic bacterium or virus. The therapeutic use of oligonucleotides represents a new paradigm for drug discovery, because oligonucleotides have not been studied before as potential drugs and they are being used to intervene in processes that have not been considered sites at which drugs might act. The affinity and the specificity of binding derive from the hybridization of two oligonucleotides and therefore are theoretically much greater than can be achieved with small molecules. Furthermore, the rational design of the nucleotide sequence of antisense oligonucleotide is much more straightforward than the design of small molecules interacting with proteins. Finally, it is possible to consider the design of antisense drugs to treat a very broad range of disorders, including those not amenable to other types of treatment.
This use of antisense oligonucleotides is still in its infancy, and the initial enthusiasm must be tempered by appropriate reservations concerning practical aspects. To be useful as a drug, an antisense oligonucleotide must be much more stable than ordinary nucleic acids, and it must be able to reach its desired site of action, the interior of a cell. Questions about the technology reduce to, can oligonucleotide analogs be created that have appropriate properties to be drugs? Specifically, what are the pharmacokinetic, pharmacological, and toxicological properties of these compounds and what are the scope and potential of the medicinal chemistry of oligonucleotides? Answers to many of these questions are available now.
2. Phosphorothioate oligodeoxynucleotides
To address the problem of stability, nonconventional oligonucleotides not susceptible to degradation or to hydrolysis by nucleases have been designed. Of the first generation of oligonucleotide analogs, the phosphorothioate class is best understood and has produced the broadest range of activities (3). Phosphorothioate oligonucleotides differ from normal in that one of the nonbridging oxygens in the phosphate group is replaced by a sulfur. The resulting compound is negatively charged, chiral at each phosphorothioate, and much more resistant to nucleases than a phosphodiester bridge.
2.1. Hybridization
The hybridization of phosphorothioate oligonucleotides to DNA and RNA has been thoroughly characterized. The melting temperature (Tm) of a phosphorothioate oligodeoxynucleotide bound to RNA is lower than for a corresponding phosphodiester oligodeoxynucleotide by approximately 0.5°C per nucleotide. Compared to RNA duplex formation, a phosphorothioate oligodeoxynucleotide has a Tm approximately 2.2°C lower per nucleotide. This means that, to be effective in vitro, phosphorothioate oligodeoxynucleotides must be relatively long, typically at least 17 to 20 nucleotides, and invasion of double-stranded regions of the target RNA is difficult (4-7).
2.1.1. Interactions with Proteins
Phosphorothioate oligonucleotides bind to proteins. These interactions can be (3) nonspecific, sequence-specific, or structure-specific, each of which may have different characteristics and effects. Nonspecific binding to a wide variety of proteins has been demonstrated, most thoroughly with serum albumin. The affinity of such interactions is low. The dissociation constant (Kd) for albumin is approximately 200 µM, about the same as for its binding of aspirin or penicillin (8-10). Phosphorothioates also interact with nucleases and DNA polymerases; they are slowly metabolized by both endo and exonucleases and are competitive inhibitors of these enzymes (11). In an RNA–DNA duplex, phosphorothioates are substrates for ribonuclease H (RNaseH) (12). At higher concentrations, phosphorothioates inhibit the enzyme, presumably by binding as a single strand (11). Again, the oligonucleotides are competitive antagonists for the DNA–RNA substrate.
Phosphorothioates are competitive inhibitors of DNA polymerase a and b with respect to the DNA template and noncompetitive inhibitors of DNA polymerases g and d (12). They are also competitive inhibitors for the reverse transcriptase of HIV and inhibit its associated RNase H activity (13, 14). They bind to the cell surface protein CD4 and to protein kinase C (15). Phosphorothioates inhibit various viral polymerases (16), and they also cause potent, nonsequence-specific inhibition of RNA splicing (17).
2.1.2. In Vivo Pharmacokinetics
Binding of phosphorothioate oligonucleotides to serum albumin and alpha-2 macroglobulin provides a repository for these drugs in the serum and prevents their rapid renal excretion. Because serum protein binding is saturable, however, intact oligomer may be found in urine with high doses (18, (19, eg, 15 to 20 mg kg–1 administered intravenously to rats. Phosphorothioate oligonucleotides are rapidly and extensively absorbed after parenteral administration, as much as 70% within 4 h (20, 21). Distribution of phosphorothioate oligonucleotides from blood after absorption or intravenous administration is extremely rapid. Distribution half-lives are less than one hour (18-20, 22). Clearance from the blood and plasma exhibits complex kinetics, with a terminal elimination half-life of 40 to 60 h in all species except man, where it may be somewhat longer (21). Phosphorothioates distribute broadly to all peripheral tissues, although no evidence for significant penetration of the blood brain barrier has been reported. Liver, kidney, bone marrow, skeletal muscle, and skin accumulate the highest amounts (20, 22). Liver accumulates the drug most rapidly (20% of a dose within 1 to 2 hours) and also eliminates it most rapidly (eg, the terminal half-life from liver is 62 h and from renal medulla 156 h). Within the kidney (23), oligonucleotides are probably filtered by the glomerulus, then reabsorbed by the proximal convoluted tubule epithelial cells, perhaps mediated by interactions with specific proteins in the brush border membranes. At relatively low doses, clearance of phosphorothioate oligonucleotides is caused primarily by metabolism (19, 20, 22), mediated by exo- and endonucleases.
2.1.3. Pharmacological Activities
Phosphorothioates also have effects inconsistent with the antisense mechanism for which they were designed. Some of these effects are sequence- or structure-specific. Others result from nonspecific interactions with proteins. These effects are particularly prominent in in vitro tests for antiviral activity, when high concentrations of cells, viruses, and oligonucleotides are often incubated together (24, 25). Human immune deficiency virus (HIV) is particularly problematic, because many oligonucleotides bind to the gp120 protein (26). Moreover, uncertainty as to the mode of action of antisense oligonucleotides is certainly not limited to antiviral or just in vitro tests (27-29). These observations indicate that, before drawing conclusions, careful analysis of dose-response curves, direct analysis of target protein or RNA, and inclusion of appropriate controls are required. In addition to interactions with proteins, other factors can contribute to unexpected results, such as overrepresented sequences of RNA and unusual structures that may be adopted by oligonucleotides (26).
A relatively large number of reports of in vivo activities of phosphorothioate oligonucleotides have now appeared documenting activities after both local and systemic administration (30). However, for only a few of these reports have sufficient studies been performed to draw relatively firm conclusions concerning the mechanism of action by directly examining target RNA levels, target protein levels, and pharmacological effects, using a wide range of control oligonucleotides and examining the effects on closely related isotypes (31-34). Thus, there is a growing body of evidence that phosphorothioate oligonucleotides induce potent systemic and local effects in vivo, suggesting highly specific effects difficult to explain via any mechanism other than antisense.
2.1.4. Toxicological Properties
Phosphorothioate oligonucleotides are not toxic in themselves, but their use is limited by side effects. In rodents, this is immune stimulation (35, 36). In monkeys, it is sporadic reductions in blood pressure associated with bradycardia, which is often associated with activation of C-5 complement involving activation of the alternative complement pathway (37). All phosphorothioate oligonucleotides tested to date induce these effects, although there are slight variations in potency depending on their sequence and/or length (36, 38, 39). A second prominent toxicological effect in the monkey is on blood clotting. The mechanisms responsible for these effects are probably very complex, but preliminary data suggest that direct interactions with thrombin are at least partially responsible (40.(
In man, the toxicological profile differs. When ISIS 2922 is administered intravitreally to patients with cytomegalovirus retinitis, the most common adverse event is anterior chamber inflammation, which is easily managed with steroids. A relatively rare and dose-related adverse event is morphological changes in the retina associated with loss in peripheral vision (41). ISIS 2105, a 20-mer phosphorothioate designed to inhibit the replication of human papilloma viruses that cause genital warts, has been administered intradermally at doses as high as 3 mg/wart weekly for three weeks. Essentially no toxicities have been observed (42).
2.1.5. Therapeutic Index
An attempt to put the toxicities and their dose-response relationships into a therapeutic context is shown in Fig. 1. This is particularly important because considerable confusion has arisen concerning the potential utility of phosphorothioate oligonucleotides for selected therapeutic purposes as a result of unsophisticated interpretation of toxicological data. As can be readily seen, the immune stimulation induced by these compounds is particularly prominent in rodents and unlikely to be dose-limiting in humans. Nor have hypotensive events in humans been observed to date. This toxicity occurs at lower doses in monkeys than in man and certainly is not dose-limiting in man. On the basis of present experience, the dose-limiting toxicity in man is likely to result from blood clotting abnormalities, associated with peak plasma concentrations well in excess of 10 µg/ml. Thus, it phosphorothioate oligonucleotides have a therapeutic index that supports their evaluation for a number of therapies.

Figure 1. Plasma concentrations of ISIS 2302 at which various activities are observed. These concentrations are those of intact ISIS 23902 and were determined by extracting and analyzing plasma by capillary zone electrophoresis.
2.1.6. Clinical Activities
Significant therapeutic benefit has been reported in patients with cytomegalovirus retinitis treated locally with fomivirsen (43). ISIS 2302 administered every other day for one month also resulted in statistically significant improvement for five to six months in patients with steroid-dependent Crohn's disease in a randomized, double-blind placebo, controlled trial (44).
2.2. Medicinal Chemistry of Oligonucleotides
The core of any rational drug discovery program is medicinal chemistry. Although the synthesis of modified nucleic acids has been a subject of interest for some time, the intense focus on the medicinal chemistry of oligonucleotides dates perhaps no more than the past five years. Modifications have been made to the base, sugar, and phosphate moieties of oligonucleotides (Fig.( 2. The subjects of medicinal chemical programs include approaches to (3) create enhanced and more selective affinities for RNA or duplex structures, the ability to cleave nucleic acid targets, enhanced nuclease stability, (4) cellular uptake and distribution, and in vivo tissue distribution, metabolism, and clearance. Arguably, the most interesting modifications to date are those that alter the sugar moiety (45) and the backbon. Modifications such as 2′ methoxyethoxy enhance affinity for RNA, potency in vivo, provide a dramatic increase in stability, and reduce the potency for blood clotting and inflammatory effects. Also of interest are a number of modifications that replace the phosphate or the entire phosphate sugar backbone. Several novel chemical classes are being evaluated in animals and will shortly be studied in man, so it seems likely that a variety of chemical classes with differing properties will be available in the near future.
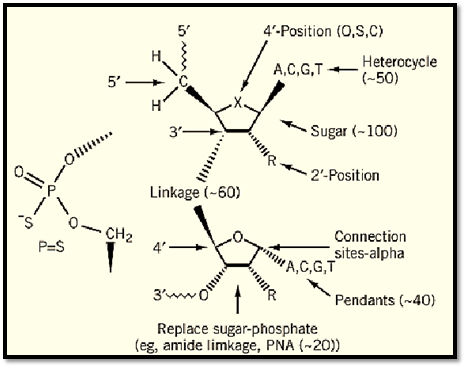
Figure 2. Isis oligonucleotide modifications.
References
1. M. Y. Chiang et al. (1991) J. Biol. Chem. 266, 18162–18171.
2. C. F. Bennett and S. T. Crooke (1996) "Oligonucleotide-based inhibitors of cytokine expression and function". In Therapeutic Modulation of Cytokines (B. Henderson, and M. W. Bodmer eds.), CRC Press, Boca Raton, pp. 171–193.
3. E. De Clercq, F. Eckstein, and T. C. Merigan (1969) Science 165, 1137–1140.
4. W. F. Lima et al. (1992) Biochemistry 31, 12055–12061.
5. T. Vickers et al. (1991) Nucleic Acids Res. 19, 3359–3368.
6. B. P. Monia et al. (1993) J. Biol. Chem. 268, 14514–14522.
7. B. P. Monia et al. (1992) J. Biol. Chem. 267, 19954–19962.
8. S. T. Crooke et al. (1996) J. Pharmacol. Exp. Ther. 277(2), 923–937.
9. R. W. Joos and W. H. Hall (1969) J. Pharmacol. Exp. Ther. 166, 113–118.
10. S. K. Srinivasan, H. K. Tewary, and P. L. Iversen (1995) Antisense Res. Dev. 5(2), 131–139.
11. S. T. Crooke et al. (1995) Biochem. J. 312(2), 599–608.
12. W. Y. Gao et al. (1992) Mol. Pharmacol. 41, 223–229.
13. C. Majumdar et al. (1989) Biochemistry 28, 1340–1346.
14. Y. Cheng, W. Gao, and F. Han (1991) Nucleosides Nucleotides 10, 155–166.
15. C. A. Stein et al. (1991) Acquired Immune Defic. Syndr. 4, 686–693.
16. C. A. Stein and Y. C. Cheng (1993) Science, 261, 1004–1012.
17. D. Hodges and S. T. Crooke (1995) Mol. Pharmacol., 48, 905–918.
18. S. Agrawal, J. Temsamani, and J. Y. Tang (1991) Proc. Natl. Acad. Sci. USA 88, 7595–7599.
19. P. Iversen (1991) Anticancer Drug Des. 6(6), 531–8.
20. P. A. Cossum et al. (1994) J. Pharmacol. Exp. Ther. 269, 89–94.
21. S. T. Crooke et al. (1994) Clin. Pharm. Ther. 56, 641–646.
22. P. A. Cossum et al. (1993) J. Pharmacol. Exp. Ther. 267, 1181–1190.
23. J. Rappaport et al. (1995) Kidney Int. 47, 1462–1469.





















