























النبات

مواضيع عامة في علم النبات
الجذور - السيقان - الأوراق
النباتات الوعائية واللاوعائية
البذور (مغطاة البذور - عاريات البذور)
الطحالب
النباتات الطبية
الحيوان
مواضيع عامة في علم الحيوان
علم التشريح
التنوع الإحيائي
البايلوجيا الخلوية
الأحياء المجهرية
البكتيريا
الفطريات
الطفيليات
الفايروسات
علم الأمراض
الاورام
الامراض الوراثية
الامراض المناعية
الامراض المدارية
اضطرابات الدورة الدموية
مواضيع عامة في علم الامراض
الحشرات
التقانة الإحيائية
مواضيع عامة في التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحيوية والميكروبات
الفعاليات الحيوية
وراثة الاحياء المجهرية
تصنيف الاحياء المجهرية
الاحياء المجهرية في الطبيعة
أيض الاجهاد
التقنية الحيوية والبيئة
التقنية الحيوية والطب
التقنية الحيوية والزراعة
التقنية الحيوية والصناعة
التقنية الحيوية والطاقة
البحار والطحالب الصغيرة
عزل البروتين
هندسة الجينات
التقنية الحياتية النانوية
مفاهيم التقنية الحيوية النانوية
التراكيب النانوية والمجاهر المستخدمة في رؤيتها
تصنيع وتخليق المواد النانوية
تطبيقات التقنية النانوية والحيوية النانوية
الرقائق والمتحسسات الحيوية
المصفوفات المجهرية وحاسوب الدنا
اللقاحات
البيئة والتلوث
علم الأجنة
اعضاء التكاثر وتشكل الاعراس
الاخصاب
التشطر
العصيبة وتشكل الجسيدات
تشكل اللواحق الجنينية
تكون المعيدة وظهور الطبقات الجنينية
مقدمة لعلم الاجنة
الأحياء الجزيئي
مواضيع عامة في الاحياء الجزيئي
علم وظائف الأعضاء
الغدد
مواضيع عامة في الغدد
الغدد الصم و هرموناتها
الجسم تحت السريري
الغدة النخامية
الغدة الكظرية
الغدة التناسلية
الغدة الدرقية والجار الدرقية
الغدة البنكرياسية
الغدة الصنوبرية
مواضيع عامة في علم وظائف الاعضاء
الخلية الحيوانية
الجهاز العصبي
أعضاء الحس
الجهاز العضلي
السوائل الجسمية
الجهاز الدوري والليمف
الجهاز التنفسي
الجهاز الهضمي
الجهاز البولي
المضادات الميكروبية
مواضيع عامة في المضادات الميكروبية
مضادات البكتيريا
مضادات الفطريات
مضادات الطفيليات
مضادات الفايروسات
علم الخلية
الوراثة
الأحياء العامة
المناعة
التحليلات المرضية
الكيمياء الحيوية
مواضيع متنوعة أخرى
الانزيمات

RNA Polymerase I Has a Bipartite Promoter
 المؤلف:
JOCELYN E. KREBS, ELLIOTT S. GOLDSTEIN and STEPHEN T. KILPATRICK
المؤلف:
JOCELYN E. KREBS, ELLIOTT S. GOLDSTEIN and STEPHEN T. KILPATRICK
 المصدر:
LEWIN’S GENES XII
المصدر:
LEWIN’S GENES XII
 الجزء والصفحة:
الجزء والصفحة:
 7-5-2021
7-5-2021
 3103
3103




+
-
20
RNA Polymerase I Has a Bipartite Promoter
KEY CONCEPTS
- The RNA polymerase I promoter consists of a core promoter and an upstream promoter element (UPE).
- The factor UBF1 wraps DNA around a protein structure to bring the core and UPE into proximity.
- SL1 includes the factor TATA-binding protein (TBP) that is involved in initiation by all three RNA polymerases.
- RNA polymerase I binds to the UBF1–SL1 complex at the core promoter.
RNA polymerase I transcribes only the genes for ribosomal RNA from a single type of promoter in a special region of the nucleus called the nucleolus. The precursor transcript includes the sequences of both large 28S and small 18S rRNAs, which are later processed by cleavages and modifications. Ribosome assembly also occurs in the nucleolus. There are many copies of the rRNA transcription unit. They alternate with nontranscribed spacers and are organized in a cluster, as discussed in the chapter titled Clusters and Repeats. The organization of the promoter, and the events involved in initiation, are illustrated in FIGURE 1. RNA polymerase I exists as a holoenzyme that contains additional factors required for initiation and is recruited by its transcription factors directly as a giant complex to the promoter.
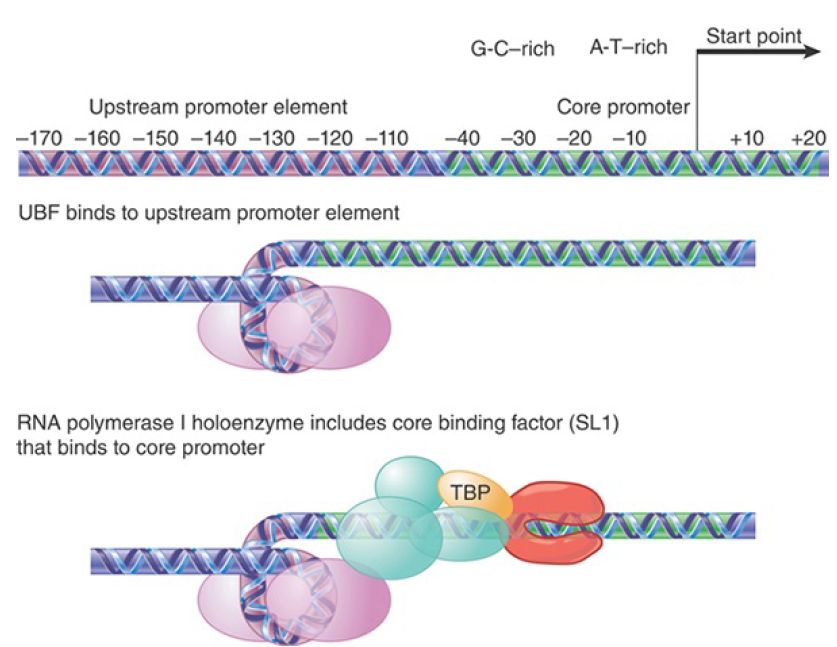
FIGURE 18.3 Transcription units for RNA polymerase I have a core promoter separated by ~70 bp from the upstream promoter element. UBF binding to the UPE increases the ability of corebinding factor to bind to the core promoter. Core-binding factor(SL 1) positions RNA polymerase I at the start point.
The promoter consists of two separate regions. The core promoter surrounds the start point, extending from −45 to +20, and is sufficient for transcription to initiate. It is generally G-C rich
(unusual for a promoter), except for the only conserved sequence element, a short A-T–rich sequence around the start point. The core promoter’s efficiency, however, is very much increased by the upstream promoter element (UPE, sometimes also called the upstream control element, or UCE). The UPE is another G-C–rich sequence related to the core promoter sequence, extending from −180 to −107. This type of organization is common to pol I promoters in many species, although the actual sequences vary widely.
RNA polymerase I requires two ancillary transcription factors to recognize the promoter sequence. The factor that binds to the core promoter is SL1 (or TIF-1B and Rib1 in different species), which consists of four protein subunits. Two of the components of SL1 are the TATA-binding protein (TBP), a factor that also is required for initiation by RNA polymerases II and III, and a second component that is homologous to the RNA polymerase II factor TF B (see the section in this chapter titled TBP Is a Universal Factor). TBP does not bind directly to G-C−rich DNA, and DNA binding is the responsibility of the other components of SL1. It is likely that TBP interacts with RNA polymerase, probably with a common subunit or a feature that has been conserved among polymerases. SL1 enables RNA polymerase I to initiate from the promoter at a low basal frequency.
SL1 has primary responsibility for RNA polymerase recruitment, proper localization of polymerase at the start point, and promoter escape. As will be discussed later, a comparable function is provided for RNA polymerases II and III by a factor that consists of TBP and other proteins. Thus, a common feature in initiation by all three polymerases is a reliance on a “positioning factor” that consists of TBP associated with proteins that are specific for each type of promoter. The exact mode of action is different for each of the TBP-dependent positioning factors; at the promoter for RNA polymerase I it does not bind DNA, whereas at TATA box–containing promoters for RNA polymerase II it is the principal means for locating the factor on DNA. For high-frequency initiation, the transcription factor UBF is required. This is a single polypeptide that binds to a G-C–rich II
element in the UPE. UBF has multiple functions. UBF is required to maintain open chromatin structure. It prevents histone HI binding, and therefore prevents assembly of inactive chromatin. It stimulates promoter release by the RNA polymerase, and it stimulates SL1. One indication of how UBF interacts with SL1 is given by the importance of the spacing between UBF and the core promoter. This can be changed by distances involving integral numbers of turns of DNA, but not by distances that introduce half turns. UBF binds to the minor groove of DNA and wraps the DNA in a loop of almost 360° turn on the protein surface, with the result that the core promoter and UPE come into close proximity, enabling UBF to stimulate binding of SL1 to the promoter.
Figure 1.shows initiation as a series of sequential interactions. RNA polymerase I, however, exists as a holoenzyme that contains most or all of the factors required for initiation, and it is probably recruited directly to the promoter. Following initiation, RNA polymerase I, like RNA polymerase II, requires a special factor, the RNA polymerase I PafI complex, for efficient elongation.
 الاكثر قراءة في مواضيع عامة في الاحياء الجزيئي
الاكثر قراءة في مواضيع عامة في الاحياء الجزيئي
 اخر الاخبار
اخر الاخبار
اخبار العتبة العباسية المقدسة

الآخبار الصحية


مواضيع ذات صلة
 قسم الشؤون الفكرية يصدر كتاباً يوثق تاريخ السدانة في العتبة العباسية المقدسة
قسم الشؤون الفكرية يصدر كتاباً يوثق تاريخ السدانة في العتبة العباسية المقدسة "المهمة".. إصدار قصصي يوثّق القصص الفائزة في مسابقة فتوى الدفاع المقدسة للقصة القصيرة
"المهمة".. إصدار قصصي يوثّق القصص الفائزة في مسابقة فتوى الدفاع المقدسة للقصة القصيرة (نوافذ).. إصدار أدبي يوثق القصص الفائزة في مسابقة الإمام العسكري (عليه السلام)
(نوافذ).. إصدار أدبي يوثق القصص الفائزة في مسابقة الإمام العسكري (عليه السلام)



















