























النبات

مواضيع عامة في علم النبات
الجذور - السيقان - الأوراق
النباتات الوعائية واللاوعائية
البذور (مغطاة البذور - عاريات البذور)
الطحالب
النباتات الطبية
الحيوان
مواضيع عامة في علم الحيوان
علم التشريح
التنوع الإحيائي
البايلوجيا الخلوية
الأحياء المجهرية
البكتيريا
الفطريات
الطفيليات
الفايروسات
علم الأمراض
الاورام
الامراض الوراثية
الامراض المناعية
الامراض المدارية
اضطرابات الدورة الدموية
مواضيع عامة في علم الامراض
الحشرات
التقانة الإحيائية
مواضيع عامة في التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحيوية والميكروبات
الفعاليات الحيوية
وراثة الاحياء المجهرية
تصنيف الاحياء المجهرية
الاحياء المجهرية في الطبيعة
أيض الاجهاد
التقنية الحيوية والبيئة
التقنية الحيوية والطب
التقنية الحيوية والزراعة
التقنية الحيوية والصناعة
التقنية الحيوية والطاقة
البحار والطحالب الصغيرة
عزل البروتين
هندسة الجينات
التقنية الحياتية النانوية
مفاهيم التقنية الحيوية النانوية
التراكيب النانوية والمجاهر المستخدمة في رؤيتها
تصنيع وتخليق المواد النانوية
تطبيقات التقنية النانوية والحيوية النانوية
الرقائق والمتحسسات الحيوية
المصفوفات المجهرية وحاسوب الدنا
اللقاحات
البيئة والتلوث
علم الأجنة
اعضاء التكاثر وتشكل الاعراس
الاخصاب
التشطر
العصيبة وتشكل الجسيدات
تشكل اللواحق الجنينية
تكون المعيدة وظهور الطبقات الجنينية
مقدمة لعلم الاجنة
الأحياء الجزيئي
مواضيع عامة في الاحياء الجزيئي
علم وظائف الأعضاء
الغدد
مواضيع عامة في الغدد
الغدد الصم و هرموناتها
الجسم تحت السريري
الغدة النخامية
الغدة الكظرية
الغدة التناسلية
الغدة الدرقية والجار الدرقية
الغدة البنكرياسية
الغدة الصنوبرية
مواضيع عامة في علم وظائف الاعضاء
الخلية الحيوانية
الجهاز العصبي
أعضاء الحس
الجهاز العضلي
السوائل الجسمية
الجهاز الدوري والليمف
الجهاز التنفسي
الجهاز الهضمي
الجهاز البولي
المضادات الميكروبية
مواضيع عامة في المضادات الميكروبية
مضادات البكتيريا
مضادات الفطريات
مضادات الطفيليات
مضادات الفايروسات
علم الخلية
الوراثة
الأحياء العامة
المناعة
التحليلات المرضية
الكيمياء الحيوية
مواضيع متنوعة أخرى
الانزيمات

Yeast Can Switch Silent and Active Mating-Type Loci
 المؤلف:
JOCELYN E. KREBS, ELLIOTT S. GOLDSTEIN and STEPHEN T. KILPATRICK
المؤلف:
JOCELYN E. KREBS, ELLIOTT S. GOLDSTEIN and STEPHEN T. KILPATRICK
 المصدر:
LEWIN’S GENES XII
المصدر:
LEWIN’S GENES XII
 الجزء والصفحة:
الجزء والصفحة:
 17-4-2021
17-4-2021
 2567
2567




+
-
20
Yeast Can Switch Silent and Active Mating-Type Loci
KEY CONCEPTS
- The yeast mating-type locus MAT has either the MATa or MATα genotype.
- Yeast with the dominant allele HO switch their mating type at a frequency of about 10-6 .
- The allele at MAT is called the active cassette.
- There are also two silent cassettes, HMLα and HMRa.
- Switching occurs if MATa is replaced by HMRα or MATα is replaced by HMRa.
The yeast S. cerevisiae can propagate in either the haploid or diploid condition. Conversion between these states takes place by mating (fusion of haploid cells to give a diploid) and by sporulation (meiosis of diploids to give haploid spores). The ability to engage in these activities is determined by the mating type of the strain, which can be either a or α. Haploid cells of type a can mate only with haploid cells of type α to generate diploid cells of type a/α. The diploid cells can sporulate to regenerate haploid spores of either type.
Mating behavior is determined by the genetic information present at the MAT locus. Cells that carry the MATa allele at this locus are type a; likewise, cells that carry the MATα allele are type α.
Recognition between cells of opposite mating type is accomplished by the secretion of pheromones: α cells secrete the small polypeptide α factor; a cells secrete a factor. A cell of one mating type carries a surface receptor for the pheromone of the opposite type. When an a cell and an α cell encounter one another, their pheromones act on their receptors to arrest the cells in the G1 phase of the cell cycle, and various morphological changes occur (including “schmooing,” in which cells elongate toward each other).
In a successful mating, the cell cycle arrest is followed by cell and nuclear fusion to produce an a/α diploid cell.
Mating is a symmetrical process that is initiated by the interaction of pheromone secreted by one cell type with the receptor carried by the other cell type. The only genes that are uniquely required for the response pathway in a particular mating type are those coding for the receptors. Either the a factor–receptor interaction or the α factor–receptor interaction switches on the same response pathway. Mutations that eliminate steps in the common pathway have the same effects in both cell types. The pathway consists of a signal transduction cascade that leads to the synthesis of products that make the necessary changes in cell morphology and gene expression for mating to occur.
Much of the information about the yeast mating-type pathway was deduced from the properties of mutations that eliminate the ability of a and/or α cells to mate. The genes identified by such mutations are called STE (for sterile). Mutations in the genes for the pheromones or receptors are specific for individual mating types, whereas mutations in the other STE genes eliminate mating in both a and α cells. This situation is explained by the fact that the events that follow the interaction of factor with receptor are identical for both types.
Some yeast strains have the remarkable ability to switch their mating types. These strains carry a dominant allele HO and change their mating type frequently—as often as once every generation. Strains with the recessive allele ho have a stable mating type, which is subject to change with a frequency of about 10−6.
The presence of HO causes the genotype of a yeast population to change. Irrespective of the initial mating type, within a very few generations large numbers of cells of both mating types are present, leading to the formation of MATa/MATα diploids that take over the population. The production of stable diploids from a haploid population can be viewed as the raison d’être for switching.
The existence of switching suggests that all cells contain the potential information needed to be either MATa or MATα but express only one type. Where does the information to change mating type come from? Two additional loci are needed for switching. HMLα is needed for switching to give a MATa type; HMRa is needed for switching to give a MATa type. These loci lie on the same chromosome that carries MAT. HML is far to the left and HMR is far to the right.
The mating-type cassette model is illustrated in FIGURE 1. It proposes that MAT has an active cassette of either type α or type a. HML and HMR have silent cassettes. In general, HML carries an α cassette, whereas HMR carries an a cassette. All cassettes carry information that encodes mating type, but only the active cassette at MAT is expressed. Mating-type switching occurs when the active cassette is replaced by information from a silent cassette. The newly installed cassette is then expressed.
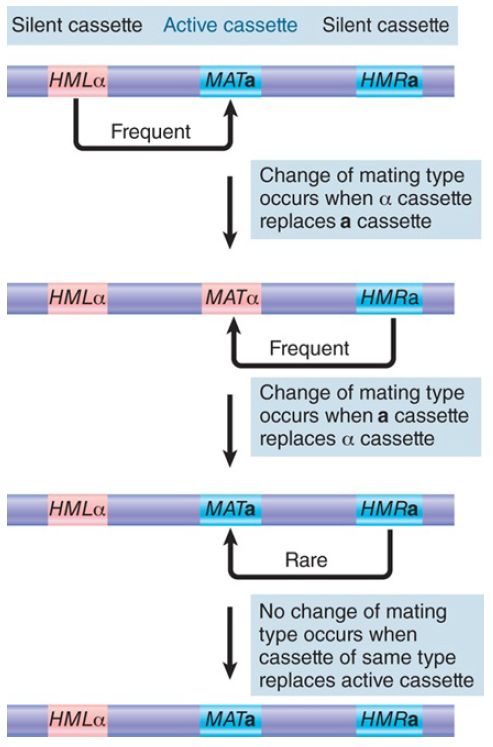
FIGURE 1. Changes of mating type occur when silent cassettes replace active cassettes of the opposite genotype; recombination occurs between cassettes of the same type, and the mating type remains unaltered.
Switching is nonreciprocal; the copy at HML or HMR replaces the allele at MAT. We know this because a mutation at MAT is lost permanently when it is replaced by switching—it does not exchange with the copy that replaces it. This is, in effect, a directed gene-conversion event. The directionality is established by the DSB initiation event, which occurs in the active MAT gene and not in the silent cassettes.
If the silent copy present at HML or HMR is mutated, switching introduces a mutant allele into the MAT locus. The mutant copy at HML or HMR remains there through an indefinite number of switches.
Mating-type switching is a directed event, in which there is only one recipient (MAT), but two potential donors (HML and HMR). Switching usually involves replacement of MATa by the copy at HMLα or replacement of MATα by the copy at HMRa. In 80% to 90% of switches, the MAT allele is replaced by one of the opposite type. This is determined by the phenotype of the cell. Cells of a phenotype preferentially choose HML as donor; cells of α phenotype preferentially choose HMR.
Several groups of genes are involved in establishing and switching mating type. In addition to the genes that directly determine mating type, they include genes needed to repress the silent cassettes, to switch mating type, or to execute the functions involved in mating, and, most important, the homologous recombination factors described earlier in this chapter.
By comparing the sequences of the two silent cassettes (HMLα and HMRa) with the sequences of the two types of active cassettes (MATa and MATα), the sequences that determine mating type can be delineated. The organization of the mating-type loci is summarized in FIGURE 2. Each cassette contains common sequences that flank a central region that differs in the a and α types of cassette (called Y a or Yα). On either side of this region, the flanking sequences are virtually identical, although they are shorter at HMR. The active cassette at MAT is transcribed from a promoter within the Y region.
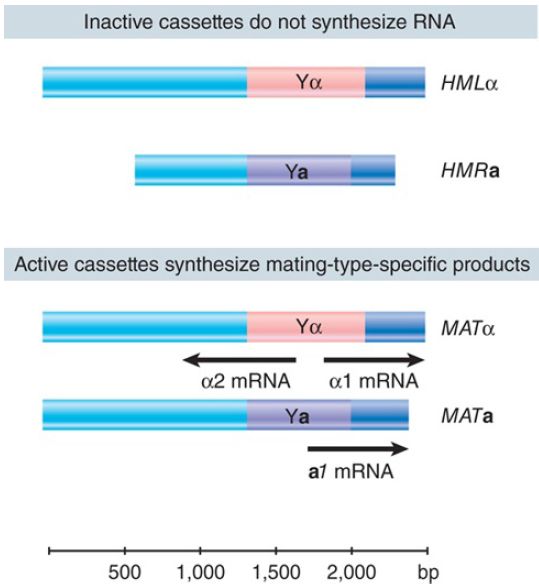
FIGURE 2.Silent cassettes have the same sequences as the corresponding active cassettes, except for the absence of the extreme flanking sequences in HMRa. Only the Y region changes
between a and α types.
 الاكثر قراءة في مواضيع عامة في الاحياء الجزيئي
الاكثر قراءة في مواضيع عامة في الاحياء الجزيئي
 اخر الاخبار
اخر الاخبار
اخبار العتبة العباسية المقدسة

الآخبار الصحية


مواضيع ذات صلة
 قسم الشؤون الفكرية يصدر كتاباً يوثق تاريخ السدانة في العتبة العباسية المقدسة
قسم الشؤون الفكرية يصدر كتاباً يوثق تاريخ السدانة في العتبة العباسية المقدسة "المهمة".. إصدار قصصي يوثّق القصص الفائزة في مسابقة فتوى الدفاع المقدسة للقصة القصيرة
"المهمة".. إصدار قصصي يوثّق القصص الفائزة في مسابقة فتوى الدفاع المقدسة للقصة القصيرة (نوافذ).. إصدار أدبي يوثق القصص الفائزة في مسابقة الإمام العسكري (عليه السلام)
(نوافذ).. إصدار أدبي يوثق القصص الفائزة في مسابقة الإمام العسكري (عليه السلام)



















