Gene Conversion Accounts for Interallelic Recombination
KEY CONCEPTS
-Heteroduplex DNA that is created by recombination can have mismatched sequences where the recombining alleles are not identical.
- Repair systems may remove mismatches by changing one of the strands so its sequence is complementary to the other.
-Mismatch repair of heteroduplex DNA generates nonreciprocal recombinant products called gene conversions.
The involvement of heteroduplex DNA explains the characteristics of recombination between alleles; indeed, allelic recombination provided the impetus for the development of a recombination model that invoked heteroduplex DNA as an intermediate. When recombination between alleles was discovered, the natural assumption was that it takes place by the same mechanism of reciprocal recombination that applies to more distant loci. That is to say, both events are initiated in the same manner: A DSB repair event can occur within a locus to generate a reciprocal pair of recombinant chromosomes. In the close quarters of a single gene,
however, formation and repair of heteroduplex DNA itself is responsible for the gene-conversion event.
Individual recombination events can be studied in the ascomycete fungi, because the products of a single meiosis are held together in a large cell called the ascus (or, less commonly, the tetrad). Even better is that in some fungi the four haploid nuclei produced by meiosis are arranged in a linear order. (Actually, a mitotic division occurs after the production of these four nuclei, giving a linear series of eight haploid nuclei.) FIGURE 1. shows that each of these nuclei effectively represents the genetic character of one of the eight strands of the four chromosomes produced by meiosis.
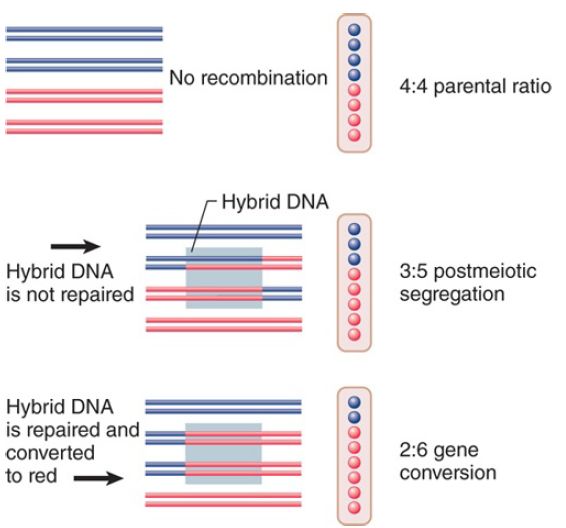
FIGURE 1. Spore formation in ascomycetes allows determination of the genetic constitution of each of the DNA strands involved in meiosis.
Meiosis in a heterozygous diploid should generate four copies of each allele in these fungi. This is seen in the majority of spores. Some spores, however, have abnormal ratios. These spores are explained by the formation and correction of heteroduplex DNA in the region in which the alleles differ. Figure 1 illustrates a recombination event in which a length of hybrid DNA occurs on one of the four meiotic chromosomes, a possible outcome of recombination initiated by a DSB.
Suppose that two alleles differ by a single point mutation. When a strand exchange occurs to generate heteroduplex DNA, the two strands of the heteroduplex will be mispaired at the site of
mutation. Thus, each strand of DNA carries different genetic information. If no change is made in the sequence, the strands separate at the ensuing replication, each giving rise to a duplex that
perpetuates its information. This event is called postmeiotic segregation, because it reflects the separation of DNA strands after meiosis. Its importance is that it demonstrates directly the existence of heteroduplex DNA in recombining alleles.
Another effect is seen when examining recombination between alleles: The proportions of the alleles differ from the initial 4:4 ratio. This effect is called gene conversion. It describes a nonreciprocal transfer of information from one chromatid to another.
Gene conversion results from exchange of strands between DNA molecules, and the change in sequence may have either of two causes at the molecular level, known as gap repair or mismatch repair:
- Gap repair: As indicated by the DSBR model , one DNA duplex may act as a donor of genetic information that directly replaces the corresponding sequences in the recipient
duplex by a process of gap generation, strand exchange, and gap filling.
- Mismatch repair: As part of the exchange process, heteroduplex DNA is generated when a single strand from one duplex pairs with its complement in the other duplex. Repair systems recognize mispaired bases in heteroduplex DNA, and then may excise and replace one of the strands to restore complementarity (see the chapter titled Repair Systems). Such an event converts the strand of DNA representing one allele into the sequence of the other allele.
Gene conversion does not depend on crossing over, but rather is correlated with it. A large proportion of the aberrant asci show genetic recombination between two markers on either side of a site of interallelic gene conversion. This is exactly what would be predicted if the aberrant ratios result from initiation of the recombination process , but with an approximately equal probability of resolving the structure with or without recombination. The implication is that fungal chromosomes initiate crossing over about twice as often as would be expected from the measured frequency of recombination between distant genes.
Various biases are seen when recombination is examined at the molecular level. Either direction of gene conversion may be equally likely, or allele-specific effects may create a preference for one direction. Gradients of recombination may fall away from hotspots.
We now know that recombination hotspots represent sites at which DSBs are preferentially initiated, and that the gradient is correlated with the extent to which the gap at the hotspot is enlarged and converted to long single-stranded ends (see the section in this chapter titled The Synaptonemal Complex Forms After Double- Strand Breaks).
Some information about the extent of gene conversion is provided by the sequences of members of gene clusters. Usually, the products of a recombination event will separate and become unavailable for analysis at the level of DNA sequence. When a chromosome carries two (nonallelic) genes that are related, though, they may recombine by an “unequal crossing-over” event . All we need to note for now is that a heteroduplex may be formed between the two nonallelic genes. Gene conversion effectively converts one of the nonallelic genes to the sequence of the other.
The presence of more than one gene copy on the same chromosome provides a footprint to trace these events. For example, if heteroduplex formation and gene conversion occurred over part of one gene, this part may have a sequence identical with, or very closely related to, the other gene, whereas the remaining part shows more divergence. Available sequences suggest that gene-conversion events may extend for considerable distances, up to a few thousand bases.





















