X Chromosomes Undergo Global Changes
KEY CONCEPTS
- One of the two X chromosomes is inactivated at random in each cell during embryogenesis of eutherian mammals.
- In exceptional cases where there are more than two X chromosomes, all but one are inactivated.
- The X-inactivation center (Xic) is a cis-acting region on the X chromosome that is necessary and sufficient to ensure that only one X chromosome remains active.
- Xic includes the Xist gene, which codes for an RNA that is found only on inactive X chromosomes.
- Xist recruits Polycomb complexes, which modify histones on the inactive X chromosome.
- Xist spreads along the X chromosome by binding to distal sites relative to the Xic.
- The mechanism that is responsible for preventing Xist RNA from accumulating on the active chromosome is unknown.
For species with chromosomal sex determination, the sex of the individual presents an interesting problem for gene regulation because of the variation in the number of X chromosomes. If Xlinked genes were expressed equally in each sex, females would have twice as much of each product as males. The importance of avoiding this situation is shown by the existence of dosage compensation, which equalizes the level of expression of X-linked genes in the two sexes. Dosage compensation mechanisms used in different species are summarized in FIGURE .1:
- In mammals, one of the two female X chromosomes is inactivated during embryogenesis. The result is that females have only one active X chromosome, which is the same situation found in males. The active X chromosome of females and the single X chromosome of males are expressed at the same level. (Note that both X chromosomes are active during early embryogenesis in females, and the inactive X chromosome actually retains about 5% activity.)
- In Drosophila, the expression of the single male X chromosome is doubled relative to the expression of each female X chromosome.
- In Caenorhabditis elegans, the expression of each female (hermaphrodite) X chromosome is halved relative to the expression of the single male X chromosome.
The common feature in all these mechanisms of dosage compensation is that the entire chromosome is the target for regulation. A global change occurs that quantitatively affects almost all of the promoters on the chromosome. Inactivation of the X chromosome in mammalian females is well documented, with the entire chromosome becoming heterochromatic.
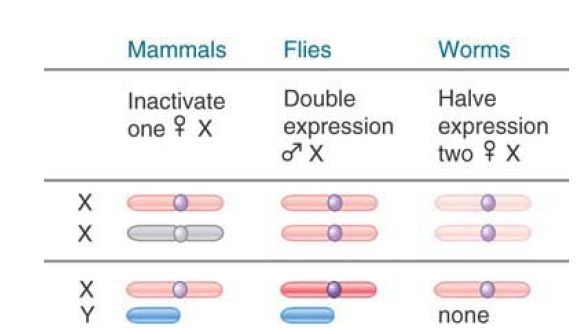
FIGURE .1 Different means of dosage compensation are used to equalize X chromosome expression in males and females.
The twin properties of heterochromatin are its condensed state and associated inactivity . It can be divided into two types:
- Constitutive heterochromatin contains specific sequences that have no coding function. These include satellite DNAs, which are often found at the centromeres. These regions are invariably heterochromatic because of their intrinsic nature.
- Facultative heterochromatin takes the form of chromosome segments or entire chromosomes that are inactive in one cell lineage, though they can be expressed in other lineages. The best example is the mammalian X chromosome. The inactive X chromosome is perpetuated in a heterochromatic state, whereas the active X chromosome is euchromatic. Either X chromosome has an equal chance of being inactivated; thus, identical DNA sequences are involved in both states. Once the inactive state has been established, it is inherited by descendant cells. This is an example of epigenetic inheritance, because it does not depend on the DNA sequence.
The basic view of the situation of the female mammalian X chromosomes was formed by the single X hypothesis in 1961. Female mice that are heterozygous for X-linked coat color mutations have a variegated phenotype in which some areas of the coat are wild type but others are mutant. FIGURE 2 shows that this can be explained if one of the two X chromosomes is inactivated at random in each cell of a small precursor population. Cells in which the X chromosome carrying the wild-type gene is inactivated give rise to progeny that express only the mutant allele on the active chromosome. Cells derived from a precursor where the other chromosome was inactivated have an active wild-type gene. In the case of coat color, cells descended from a particular precursor stay together and thus form a patch of the same color, creating the pattern of visible variegation (calico cats are a familiar example of this phenomenon). In other cases, individual cells in a population will express one or the other of X-linked alleles; for example, in heterozygotes for the X-linked locus G6PD, any particular red blood cell will express only one of the two allelic forms. (Random inactivation of one X chromosome occurs in eutherian mammals. In marsupials, the choice is directed: It is always the X chromosome inherited from the father that is inactivated.)
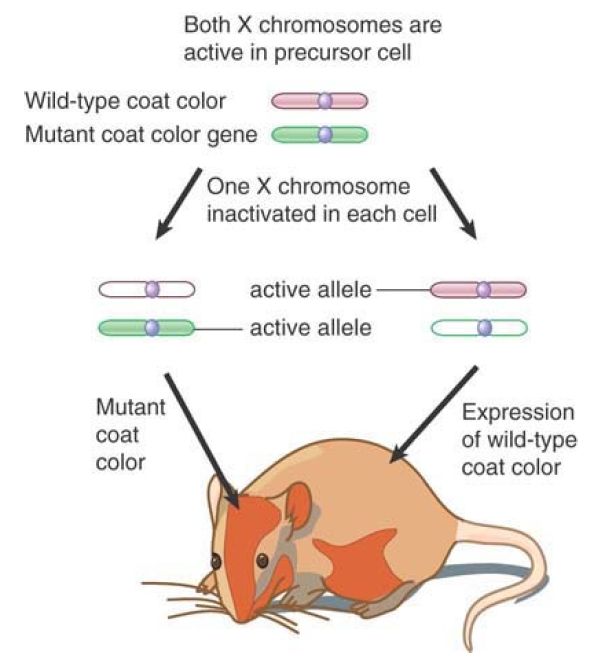
FIGURE 2. X-linked variegation is caused by the random inactivation of one X chromosome in each precursor cell. Cells in which the wild-type allele (pink) is on the active chromosome have the wild-type phenotype; cells in which the mutant allele (green) is on the active chromosome have the mutant phenotype.
Inactivation of the X chromosome in females is governed by the n –1 rule: Regardless of how many X chromosomes are present, all but one will be inactivated. Normal females of course have two X chromosomes, but in rare cases where nondisjunction has generated a genotype of three or more X chromosomes, only one X chromosome remains active. This suggests a general model in which a specific event is limited to one X chromosome that protects it from an inactivation mechanism that applies to all the others.
A single locus on the X chromosome is sufficient for inactivation. When a translocation occurs between the X chromosome and an autosome, this locus is present on only one of the reciprocal products, and only that product can be inactivated. By comparing different translocations, it is possible to map this locus, which is called the Xic (X-inactivation center). A cloned region of 450 kb contains all the properties of the Xic. When this sequence is inserted as a transgene onto an autosome, the autosome becomes subject to inactivation (at least in a cell culture system). Pairing of Xic loci on the two X chromosomes has been implicated in the mechanism for the random choice of X-inactivation. Moreover, differences in sister chromatid cohesion correlates with the outcome of the choice of the X chromosome to be inactivated, indicating that alternate states present before the inactivation process may direct the choice of which X chromosome will become inactivated.
Xic is a cis-acting locus that contains the information necessary to count X chromosomes and inactivate all copies but one. Inactivation spreads from Xic along the entire X chromosome. When Xic is present on an X chromosome–autosome translocation, inactivation spreads into the autosomal regions (although the effect is not always complete).
Xic is a complex genetic locus that expresses several long noncoding RNAs (ncRNAs). The most important of these is a gene called Xist (X inactive-specific transcript), which is stably expressed only on the inactive X chromosome. The behavior of this gene is effectively the opposite of all other loci on the chromosome, which are turned off. Deletion of Xist prevents an X chromosome from being inactivated. It does not, however, interfere with the counting mechanism (because other X chromosomes can be inactivated). Thus, we can distinguish two features of Xic: (1) an unidentified element(s) required for counting and (2) the Xist gene required for inactivation.
The n – 1 rule suggests that stabilization of Xist RNA is the “default” and that some blocking mechanism prevents stabilization at one X chromosome (which will be the active X chromosome). This means that even though Xic is necessary and sufficient for a chromosome to be inactivated, the products of other loci are necessary for the establishment of an active X chromosome.
The Xist transcript is regulated in a negative manner by Tsix, its antisense partner. Loss of Tsix expression on the future inactive X chromosome permits Xist to become upregulated and stabilized, and persistence of Tsix on the future active X chromosome prevents Xist upregulation. Tsix is, in turn, regulated by Xite, which has a Tsix-specific enhancer and is located 10 kb upstream of Tsix. FIGURE .3 illustrates the role of Xist RNA in X-inactivation. Xist codes for an ncRNA that lacks open reading frames. The Xist RNA “coats” the X chromosome from which it is synthesized, which suggests that it has a structural role. Prior to X-inactivation, it is synthesized by both female X chromosomes. Following inactivation, the RNA is found only on the inactive X chromosome. The transcription rate remains the same before and after inactivation, so the transition depends on posttranscriptional events.
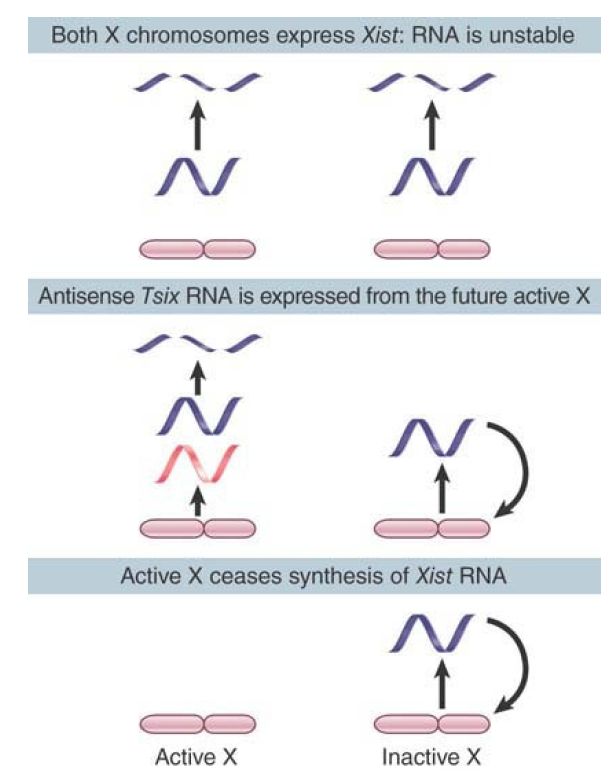
FIGURE .3 X-inactivation involves stabilization of Xist RNA, which coats the inactive chromosome. Tsix prevents Xist expression on the future active X chromosome.
Prior to X-inactivation, Xist RNA decays with a half-life of approximately 2 hours. X-inactivation is mediated by stabilizing the Xist RNA on the inactive X chromosome. The Xist RNA shows a punctate distribution along the X chromosome, which suggests that association with proteins to form particulate structures may be the means of stabilization. Xist spreads along the X chromosome beginning at the Xic and moves distally to silence regions of the X chromosome. It is not yet known what other factors may be involved in this reaction or how the Xist RNA is limited to spreading in cis along the chromosome.
Accumulation of Xist on the future inactive X chromosome results in exclusion of transcription machinery (such as RNA polymerase II) and leads to the recruitment of Polycomb repressor complexes (PRC1 and PRC2), which trigger a series of chromosome-wide histone modifications (H2AK119 ubiquitination, H3K27 methylation, H4K20 methylation, and H4 deacetylation). Late in the process, an inactive X-specific histone variant, macroH2A, is incorporated into the chromatin, and promoter DNA is methylated, resulting in gene silencing. These changes are shown in FIGURE .4. At this point, the heterochromatic state of the inactive X is stable, and Xist is not required to maintain the silent state of the chromosome.
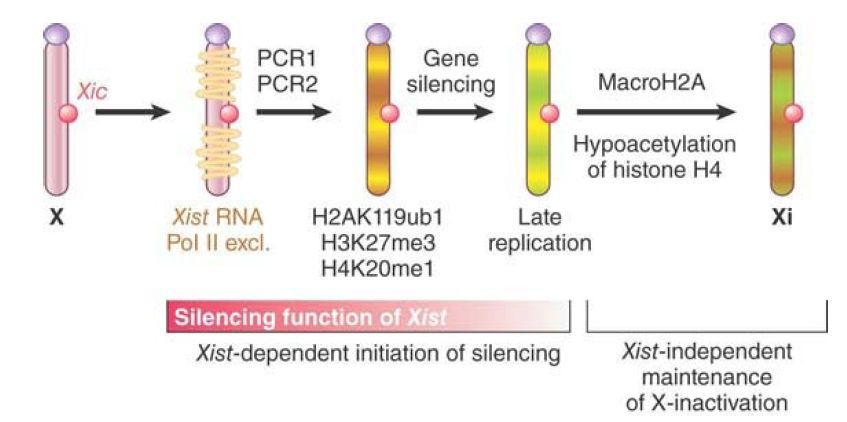
FIGURE .4 Xist RNA produced from the Xic locus accumulates on the future inactive X chromosome (Xi). This excludes transcription machinery, such as RNA polymerase II (Pol II).
Polycomb group complexes are recruited to the Xist-covered chromosome and establish chromosome-wide histone modifications. Histone macroH2A becomes enriched on the Xi, and promoters of genes on the Xi are methylated. In this phase Xinactivation is irreversible and Xist is not required for maintenance of the silent state.
Data from A. Wutz and J. Gribnau, Curr. Opin. Genet. Dev. 17 (2007): 387–393.
Despite these findings, none of the chromatin components or modifications found have been shown on their own to be essential for X chromosome silencing, indicating potential redundancy among them or the existence of pathways that have yet to be identified. Global changes also occur in other types of dosage compensation. In Drosophila, a large ribonucleoprotein complex, MSL, is found only in males, where it localizes to the X chromosome. This complex contains two noncoding RNAs, which appear to be needed for localization to the male X chromosome (perhaps analogous to the localization of Xist to the inactive mammalian X chromosome), and a histone acetyltransferase that acetylates histone H4 on K16 throughout the male X chromosome. The net result of the action of this complex is the twofold increase in transcription of all genes on the male X chromosome. The next section presents a third mechanism for dosage compensation, a global reduction in X-linked gene expression in XX (hermaphrodite) nematodes.





















