Histone Phosphorylation Affects Chromatin Structure
Key concept
- Histone phosphorylation is linked to transcription, repair, chromosome condensation, and cell-cycle progression.
All histones can be phosphorylated in vivo in different contexts. Histones are phosphorylated in three circumstances:
- Cyclically during the cell cycle
- In association with chromatin remodeling during transcription
- During DNA repair
It has long been known that the linker histone H1 is phosphorylated at mitosis, and H1 is an extremely good substrate for the Cdc2 kinase that controls cell division. This led to speculation that the phosphorylation might be connected with the condensation of chromatin, but so far no direct effect of this phosphorylation event has been demonstrated, and it is not known whether it plays a role in cell division. In Tetrahymena, it is possible to delete all the genes for H1 without significantly affecting the overall properties of chromatin, resulting in a relatively small effect on the ability of chromatin to condense at mitosis. Some genes are activated and others are repressed by this change, which suggests that there are alterations in local structure. Mutations that eliminate sites of phosphorylation in H1 have no effect, but mutations that mimic the effects of phosphorylation produce a phenotype that resembles the deletion. This suggests that the effect of phosphorylating H1 is to eliminate its effects on local chromatin structure.
Phosphorylation of serine 10 of histone H3 is linked to transcriptional activation (where it promotes acetylation of K14 in the same tail) and to chromosome condensation and mitotic progression. In Drosophila melanogaster, loss of a kinase that phosphorylates histone H3S10 (JIL-1) has devastating effects on chromatin structure. FIGURE 1compares the usual extended structure of the polytene chromosome (upper photograph) with the structure that is found in a null mutant that has no JIL-1 kinase (lower photograph). The absence of JIL-1 is lethal, but the chromosomes can be visualized in the larvae before they die.
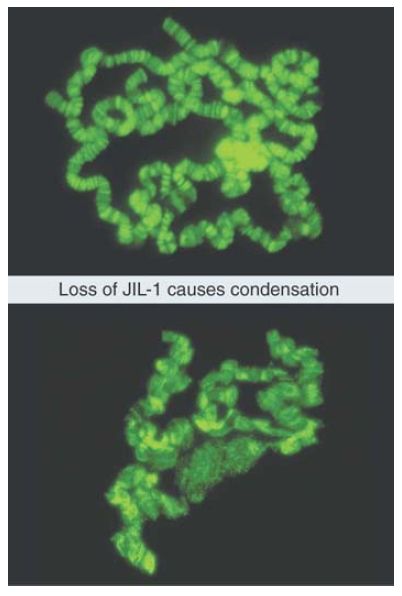
FIGURE 1. Flies that have no JIL-1 kinase have abnormal polytene chromosomes that are condensed instead of extended.
Photos courtesy of Jorgen Johansen and Kristen M. Johansen, Iowa State University.
This suggests that H3 phosphorylation is required to generate the more extended chromosome structure of euchromatic regions. JIL-1 also associates with the complex of proteins that binds to the X chromosome to increase its gene expression in males and JIL-1–dependent H3S10 phosphorylation also antagonizes H3K9 dimethylation, a heterochromatic mark. These results are consistent with a role for JIL-1 in promoting an active chromatin conformation. Interestingly, H3S10 phosphorylation by JIL-1 is itself promoted by acetylation of H4K12 by the ATAC acetyltransferase complex; these complicated interactions make it challenging to determine whether one single modification is key for the transitions in chromatin structure or whether several modifications must occur together. It is also not clear how this role of H3 phosphorylation in promoting transcriptionally active chromatin is related to the requirement for H3 phosphorylation to initiate chromosome condensation in at least some species (including mammals and the ciliate Tetrahymena).
This results in somewhat conflicting impressions of the roles of histone phosphorylation. Where it is important in the cell cycle, it is likely to be as a signal for condensation. Its effect in transcription and repair appears to be the opposite, where it contributes to open chromatin structures compatible with transcription activation and repair processes.It is possible, of course, that phosphorylation of different histones, or even of different amino acid residues in one histone, has opposite effects on chromatin structure.





















