RNA Polymerase III Uses Downstream and Upstream Promoters
KEY CONCEPTS
- RNA polymerase III uses two types of promoters.
- Internal promoters have short consensus sequences located within the transcription unit and cause initiation to occur at a fixed distance upstream.
- Upstream promoters contain three short consensus sequences upstream of the start point that are bound by transcription factors.
- TFIII A and TFIII C bind to the consensus sequences and enable TFIII B to bind at the start point.
- TFIII B has TBP as one subunit and enables RNA polymerase to bind.
Recognition of promoters by RNA polymerase III strikingly illustrates the relative roles of transcription factors and the polymerase enzyme. The promoters fall into three general classes that are recognized in different ways by different groups of factors.
The promoters for classes I and II, 5S and tRNA genes, are internal; they lie downstream of the start point. The promoters for class III snRNA (small nuclear RNA) genes lie upstream of the start point in the more conventional manner of other promoters. In both internal and external promoters, the individual elements that are necessary for promoter function consist exclusively of sequences recognized by transcription factors, which, in turn, direct the binding of RNA polymerase.
The structures of the three types of promoters for RNA polymerase III are summarized in FIGURE 1Two of the promotor types are internal promoters. Each contains a bipartite structure, in which two short sequence elements are separated by a variable sequence. The 5S ribosomal gene type 1 promoter consists of a boxA sequence separated by an intermediate element (IE) from a boxC sequence; the entire boxA-IE-boxC region is often referred to as the 5S internal control region (ICR). In yeast, only the boxC element is required for transcription. The tRNA type 2 promoter consists of a boxA sequence separated from a boxB sequence. A common group of type 3 promoters encoding other small RNAs have three sequence elements that are all located upstream of the start point; these same elements are also present in a number of RNA polymerase II promoters.
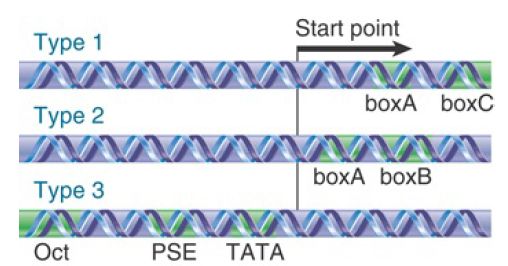
FIGURE 1. Promoters for RNA polymerase III may consist of bipartite sequences downstream of the start point, with boxA separated from either boxC or boxB, or they may consist of separated sequences upstream of the start point (Oct, PSE, TATA).
The detailed interactions are different at the two types of internal promoter, but the principle is the same. TFIII C binds downstream of the start point, either independently (tRNA type 2 promoters) or in conjunction with TF A (5S type 1 promoters). The presence of TFIII C enables the positioning factor TFIII B to bind at the start point. RNA polymerase III is then recruited.
FIGURE 2 summarizes the stages of reaction at type 2 internal promoters used for tRNA genes. The distance between boxA and boxB can vary because many tRNA genes contain a small intron. TFIII C binds to both boxA and boxB. This enables TFIII B to bind at the start point. At this point RNA polymerase III can bind.
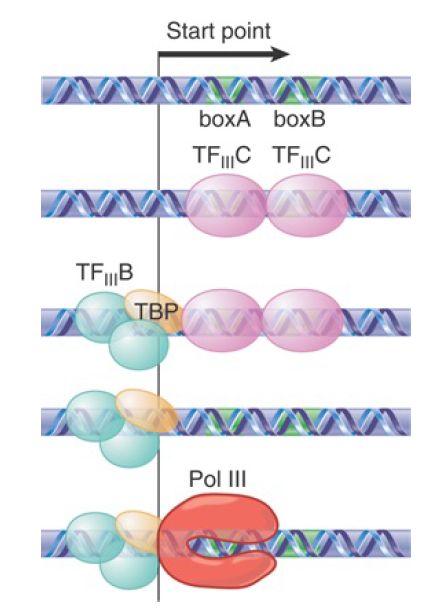
FIGURE 2. Internal type 2 pol III promoters use binding of TFIII C to boxA and boxB sequences to recruit the positioning factor TF IIIB, which recruits RNA polymerase III.
The difference at type 1 internal promoters (for 5S genes) is that TFIII A must bind at boxA to enable TFIII C to bind at boxC. TFIII A is a 5S sequence-specific binding factor that binds to the promoter and to the 5S RNA as a chaperone and gene regulator. FIGURE 18.6 shows that once TFIII C has bound events follow the same course as at type 2 promoters, with TFIII B (which contains the ubiquitous TBP) binding at the start point and RNA polymerase III joining the complex. Type 1 promoters are found only in the genes for 5S rRNA.

FIGURE 3. Internal type 1 pol III promoters use the assembly factors TFIII A and TFIII C, at boxA and boxC, to recruit the positioning factor TFIII B, which recruits RNA polymerase III.
TFIII A and TFIII C are assembly factors, whose sole role is to assist the binding of the positioning factor TFIII B at the correct location. Once TFIII B has bound, TFIII A and TFIII C can be removed from the promoter without affecting the initiation reaction. TFIII B remains bound in the vicinity of the start point, and its presence is sufficient to allow RNA polymerase III to identify and bind at the start point. Thus, TFIII B is the only true initiation factor required by RNA polymerase III. This sequence of events explains how the promoter boxes downstream can cause RNA polymerase to bind at the start point, farther upstream. Although the ability to transcribe these genes is conferred by the internal promoter, changes in the region immediately upstream of the start point can alter the efficiency of transcription.
TFIII C is a large protein complex (more than 500 kD), which is comparable in size to RNA polymerase itself, and contains six subunits. TFIII A is a member of an interesting class of proteins containing a nucleic acid–binding motif called a zinc finger. The positioning factor TFIII B consists of three subunits. It includes the same protein factor TBP that is present in the core-binding factor SL1 used for pol I promoters and in the corresponding transcription factor TF D used by RNA polymerase II. It also contains Brf, which is related to the transcription factor TFIII B that is used by RNA polymerase II and to a subunit in the RNA polymerase ISL1 factor. The third subunit is called B99; it is dispensable if the DNA duplex is partially melted, which suggests that its function is to initiate the transcription bubble. The role of B99 may be comparable to the role played by sigma factor in bacterial RNA polymerase .
The upstream region has a conventional role in the third class of polymerase III promoters. The example shown in Figure 18.4 has three upstream elements. These elements are also found in
promoters for snRNA genes that are transcribed by RNA polymerase II. (Genes for some snRNAs are transcribed by RNA polymerase II, whereas others are transcribed by RNA polymerase III.) The upstream elements function in a similar manner in promoters for both RNA polymerases II and III.
Initiation at an upstream promoter for class III RNA polymerase III can occur on a short region that immediately precedes the start point and contains only the TATA element. Efficiency of transcription, however, is much increased by the presence of the enhancer proximal sequence element (PSE) and OCT (so named because it has an 8-bp binding sequence) elements. The factors that bind at these elements interact cooperatively. The PSE element may be essential at promoters used by RNA polymerase II, whereas it is stimulatory in promoters used by RNA polymerase III.
The TATA element confers specificity for the type of polymerase (II or III) that is recognized by an snRNA promoter. It is bound by a factor that includes TBP, which actually recognizes the sequence in DNA. TBP is associated with other proteins, which are specific for the type of promoter. The function of TBP and its associated proteins is to position the RNA polymerase correctly at the start point. This is described in more detail later in the sections on RNA polymerase II.
The factors work in the same way for both types of promoters for RNA polymerase III. The factors bind at the promoter before RNA polymerase itself can bind. They form a preinitiation complex that directs binding of the RNA polymerase. RNA polymerase III does not itself recognize the promoter sequence, but binds adjacent to factors that are themselves bound just upstream of the start point. For the type I and type II internal promoters, the assembly factors ensure that TFIII B (which includes TBP) is bound just upstream of the start point, thereby providing the positioning information. For the upstream promoters, TFIII B binds directly to the region including the TATA box. This means that, irrespective of the location of the promoter sequences, factor(s) are bound close to the start point in order to direct binding of RNA polymerase III. In all cases, the chromatin must be modified and in an open configuration.





















