How Does Rho Factor Work ?
KEY CONCEPT
- Rho factor is a termination protein that binds to nascent RNA and tracks along the RNA to interact with RNA polymerase and release it from the elongation complex.
Rho factor is an essential protein in E. coli that causes transcription termination. The rho concentration may be as high as about 10% the concentration of RNA polymerase. Rho-independent termination accounts for almost half of E. coli terminators. FIGURE 1 illustrates a model for rho function. First, it binds to a sequence within the transcript upstream of the site of termination. This sequence is called a rut site (an acronym for rho utilization). The rho factor then tracks along the RNA until it catches up to RNA polymerase. When the RNA polymerase reaches the termination site, rho first freezes the structure of the polymerase and then invades the exit channel to destabilize the enzyme, causing it to release the RNA. Pausing by the polymerase at the site of termination allows time for rho factor to translocate to the hybrid stretch and is an important feature of termination.
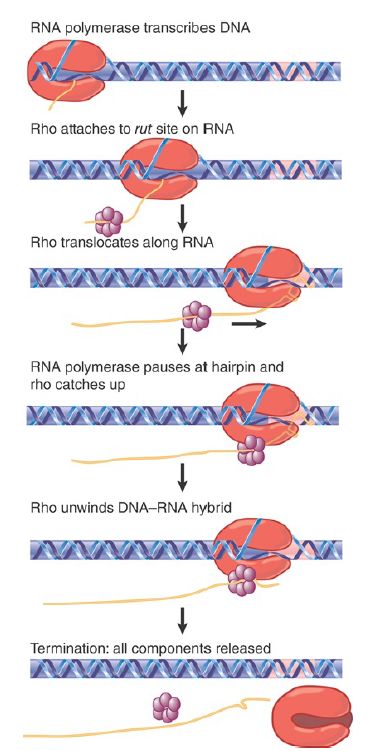
FIGURE 17.29 Rho factor binds to RNA at a rut site and translocates along RNA until it reaches the RNA–DNA hybrid in RNA polymerase, where it releases the RNA from the DNA.
We see an important general principle here. When we know the site on DNA at which some protein exercises its effect, we cannot assume that this coincides with the DNA sequence that it initially recognizes. They can be separate, and there need not be a fixed relationship between them. In fact, rut sites in different transcription units are found at varying distances preceding the sites of termination. A similar distinction is made by antitermination factors .
What actually constitutes a rut site is somewhat unclear. The common feature of rut sites is that the sequence is rich in C residues and poor in G residues and has no secondary structure. An example is given in FIGURE 2. C is by far the most common base (41%), and G is the least common base (14%). The length of rut sites also vary. As a general rule, the efficiency of a rut site increases with the length of the C-rich/G-poor region.
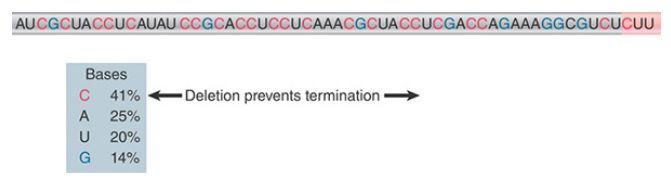
FIGURE 2. A rut site has a sequence rich in C and poor in G preceding the actual site(s) of termination. The sequence corresponds to the 3′ end of the RNA.
Rho is a member of the family of hexameric ATP-dependent helicases. Each subunit has an RNA-binding domain and an ATP hydrolysis domain. The hexamer functions by passing nucleic acid through the hole in the middle of the assembly formed from the RNA-binding domains of the subunits (FIGURE 3). The structure of rho gives some hints about how it might function. It winds RNA from the 3′ end around the exterior of the N-terminal domains, and pushes the 5′ end of the bound region into the interior, where it is bound by a secondary RNA-binding domain in the C-terminal domains. The initial form of rho is a gapped ring, but binding of the RNA converts it to a closed ring.
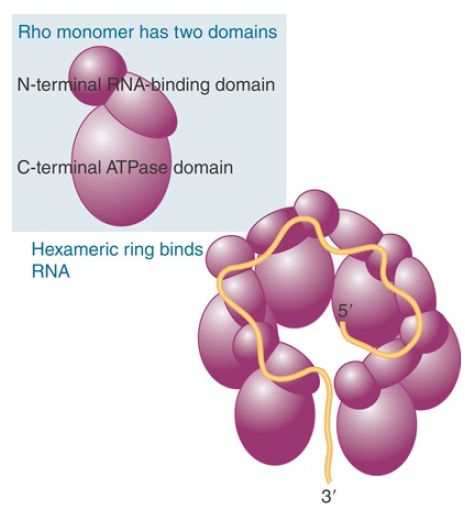
FIGURE 3 . Rho has an N-terminal, RNA-binding domain and a C-terminal ATPase domain. A hexamer in the form of a gapped ring binds RNA along the exterior of the N-terminal domains. The 5′ end of the RNA is bound by a secondary binding site in the interior of the hexamer.
After binding to the rut site, rho uses its helicase activity, driven by ATP hydrolysis, to translocate along RNA until it reaches the RNA polymerase. It then may utilize its helicase activity to unwind the duplex structure and/or interact with RNA polymerase to help release RNA.
Rho needs to translocate along RNA from the rut site to the actual point of termination. This requires the factor to move faster than RNA polymerase. The enzyme pauses when it reaches a terminator, and termination occurs if rho catches it there. Pausing is therefore important in rho-dependent termination, just as in intrinsic termination, because it gives time for the other necessary events to occur.
The coupling between transcription and translation, unique to bacteria, has important consequences for rho action. Rho must first have access to RNA upstream of the transcription complex and then moves along the RNA to catch up with RNA polymerase. As a result, its activity is impeded when ribosomes are translating an mRNA. This model explains a phenomenon that puzzled early bacterial geneticists. In some cases, a nonsense mutation in one
gene of a polycistronic transcription unit was found to prevent the expression of subsequent genes in the unit even though both genes had their own ribosome binding sites, an effect called polarity.
Rho-dependent termination sites within a transcription unit are usually masked by translating ribosomes (FIGURE 4), and therefore rho cannot act on downstream RNA polymerases. Nonsense mutations (forming stop codons) release ribosomes within the RNA of a multigene operon, though, enabling rho to terminate transcription prematurely and prevent expression of distal genes in the transcription unit even though their open reading frames contained wild-type sequences.
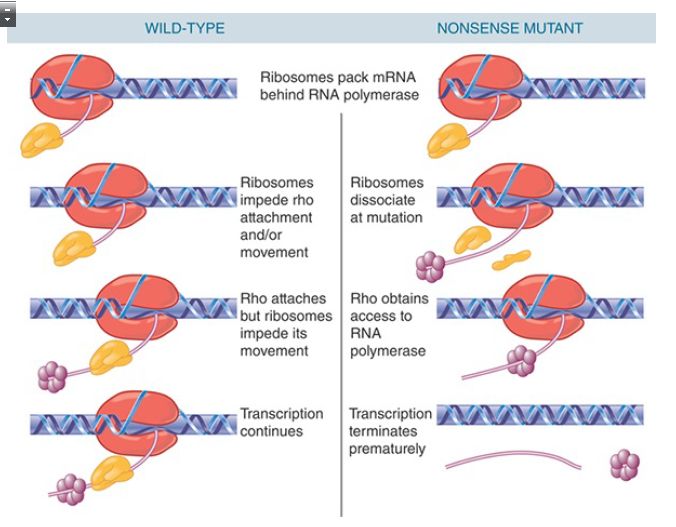
FIGURE 4. The action of rho factor may create a link between transcription and translation when a rho-dependent terminator lies soon after a nonsense mutation.
Why are stable RNAs (rRNAs and tRNAs) not subject to polarity? tRNAs are short and form extensive secondary structures that probably prevent rho binding. Parts of rRNAs also have extensive structure, but rRNAs are much longer than tRNAs, leaving ample opportunity for rho action. Cells have evolved another mechanism for preventing premature termination of rRNA transcripts, though: Proteins bind to so-called nut sites in the leader regions of the 16S/23S rRNA transcripts, forming antitermination complexes that inhibit the action of rho.
rho mutations show wide variations in their influence on termination. The basic nature of the effect is a failure to terminate. The magnitude of the failure, however, as seen in the percent of
readthrough in vivo, depends on the particular target locus. Similarly, the need for rho factor in vitro is variable. Some (rhodependent) terminators require relatively high concentrations of rho, whereas others function just as well at lower levels. This suggests that different terminators require different levels of rho factor for termination and therefore respond differently to the residual levels of rho factor in the mutants (rho mutants are usually leaky).
Some rho mutations can be suppressed by mutations in other genes. This approach provides an excellent way to identify proteins that interact with rho. The β subunit of RNA polymerase is
implicated by two types of mutation. First, mutations in the rpoB gene can reduce termination at a rho-dependent site. Second, mutations in rpoB can restore the ability to terminate transcription at rho-dependent sites in rho-mutant bacteria. It is not known, however, what function the interaction plays.





















