Introduction to TheTransposable Elements and Retroviruses
A major cause of variation in nearly all genomes is provided by transposable elements, or transposons. These are discrete sequences in the genome that are mobile; that is, they are able to transport themselves to other locations within the genome. The mark of a transposon is that it does not utilize an independent form of the element (such as phage or plasmid DNA), but rather moves directly from one site in the genome to another. Unlike most other processes involved in genome restructuring, transposition does not rely on any relationship between the sequences at the donor and recipient sites. Transposons are restricted to moving themselves, and sometimes additional sequences, to new sites elsewhere within the same genome; they are, therefore, an internal counterpart to the vectors that can transport sequences from one genome to another. They can be a major source of mutations in the genome, as shown in FIGURE 1, and have had a significant impact on the overall size of many genomes, including our own, about half of which consist of transposable elements. Transposon content in eukaryotes varies over a wide range, from 4% in yeast to 70% or more in some amphibians and plants. Plants are
particularly rich in these elements; for example, in Zea mays (maize) transposable elements make up 85% of the genome.
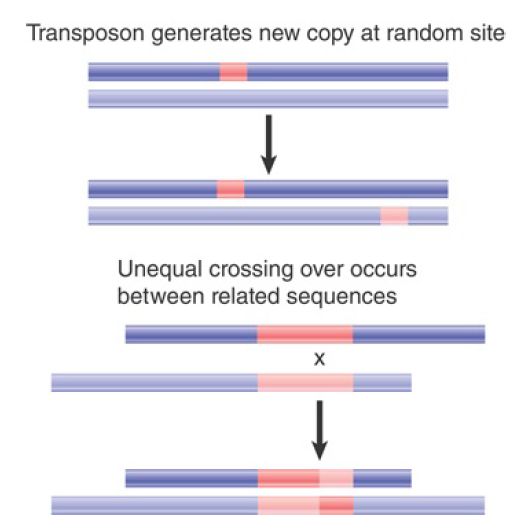
FIGURE .1 A major cause of sequence change within a genome is the movement of a transposon to a new site. This may have direct consequences on gene expression. Further, unequal crossing over between related sequences causes rearrangements. Copies of transposons can provide targets for such events.
Transposons fall into two general classes: (1) those that are able to directly manipulate DNA so as to propagate themselves within the genome (class II elements, or DNA-type elements) and (2) those whose source of mobility is the ability to make DNA copies of their RNA transcripts, which are then integrated at new sites in the genome (class I elements, or retroelements).
Transposons that mobilize via DNA are widespread in both prokaryotes and eukaryotes. Each transposon carries gene(s) that encode the enzyme activities required for its own transposition, although it may also require ancillary products of the genome in which it resides (such as DNA polymerase or DNA gyrase).
Transposition that involves an obligatory intermediate of RNA is primarily confined to eukaryotes. Transposons that employ an RNA intermediate all use some form of reverse transcriptase to translate RNA into DNA. Some of these elements are closely related to retroviral proviruses in their general organization and mechanism of transposition. As a class, these elements are called long terminal repeat (LTR) retrotransposons, or simply retrotransposons.
Members of a second class of elements that also use reverse transcriptase but lack LTRs, and that employ a distinct mode of transposition, are referred to as non-LTR retrotransposons, or simply retroposons. (The nomenclature of transposable elements is somewhat confusing in the literature, but this system of distinguishing elements by the presence or absence of the LTR reflects the modern understanding of both the evolution and the transposition mechanisms of these elements.)
Like any other reproductive cycle, the cycle of a retrovirus or retrotransposon is continuous; it is arbitrary to consider the point at which we interrupt it a “beginning.” Our perspectives of these elements are biased, though, by the forms in which we usually observe them. The interlinked cycles of retroviruses and retrotransposons are depicted in FIGURE .2. Retroviruses were first observed as infectious virus particles that were capable of transmission between cells, and so the intracellular cycle (involving duplex DNA) is thought of as the means of reproducing the RNA virus. Retrotransposons were discovered as components of the genome, and the RNA forms have been mostly characterized for their functions as mRNAs and transposition intermediates. Thus, we think of retrotransposons as genomic (duplex DNA) sequences and retroviruses as RNA–protein complexes, but this obscures the close relationship between these elements. Indeed, recent phylogenetic evidence suggests that retroviruses as a class are simply retrotransposons that have acquired envelope proteins, the inverse of the previously assumed relationship.
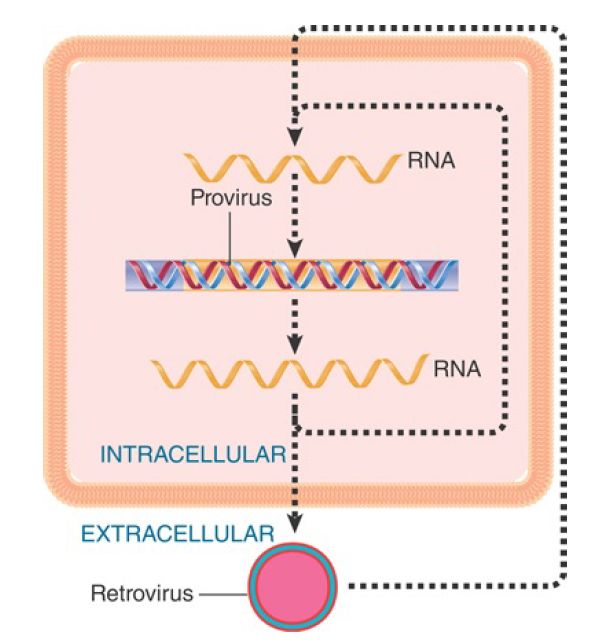
FIGURE .2 The reproductive cycles of retroviruses and retrotransposons alternate reverse transcription from RNA to DNA with transcription from DNA to RNA. Only retroviruses can generate infectious particles. Retrotransposons are confined to an intracellular cycle.
A genome may contain both functional and nonfunctional (defective) elements of either class of element. In most cases the majority of elements in a eukaryotic genome are defective and have lost the ability to transpose independently, although they may still be recognized as substrates for transposition by the enzymes produced by functional transposons. A eukaryotic genome contains a large number and variety of transposons. The relatively small fly genome has 1,572 identified transposons belonging to 96 distinct families. Larger genomes, such as those of maize and humans, can harbor hundreds of thousands of transposons. Each of these species has a genome composed of 50% to 85% transposons.
Transposable elements of all kinds can promote rearrangements of the genome directly or indirectly:
- The transposition event itself may cause deletions or inversions or lead to the movement of a host sequence to a new location.
- Transposons serve as substrates for cellular recombination systems by functioning as “portable regions of homology”; two copies of a transposon at different locations (even on different chromosomes) may provide sites for aberrant reciprocal recombination. Such exchanges result in deletions, insertions, inversions, or translocations.
The intermittent activities of a transposon seem to provide a somewhat nebulous target for natural selection. This view has prompted suggestions that most transposable elements confer neither advantage nor disadvantage on the phenotype, but could constitute “selfish DNA”—DNA concerned only with its own propagation. Indeed, in considering transposition as an event that is distinct from other cellular recombination systems we tacitly accept the view that the transposon is an independent entity that resides in the genome.
Such a relationship of the transposon to the genome would resemble that of a parasite with its host. Presumably the propagation of an element by transposition is balanced by the harm done if a transposition event inactivates a necessary gene or if the number of transposons becomes a burden on cellular systems. Yet we must remember that any transposition event conferring a selective advantage—for example, a genetic rearrangement—will lead to preferential survival of the genome carrying the active transposon.





















