Eukaryotic Nucleotide Excision Repair Pathways
KEY CONCEPTS
- Xeroderma pigmentosum (XP) is a human disease caused by mutations in any one of several nucleotide excision repair genes.
- Numerous proteins, including XP products and the transcription factor TF H, are involved in eukaryotic nucleotide excision repair.
- Global genome repair recognizes damage anywhere in the genome.
- Transcriptionally active genes are preferentially repaired via transcription-coupled repair.
- Global genome repair and transcription-coupled repair differ in their mechanisms of damage recognition (XPC vs. RNA polymerase II).
- TF II provides the link to a complex of repair enzymes.
- Mutations in the XPD component of TF II cause three different human diseases.
The general principle of excision repair in eukaryotic cells is similar to that of bacteria. Bulky lesions, such as those created by UV damage, crosslinking agents, and numerous chemical carcinogens, are also recognized and repaired by a nucleotide excision repair system. The critical role of mammalian nucleotide excision repair is seen in certain human hereditary disorders. A well-characterized example is xeroderma pigmentosum (XP), a recessive disease resulting in hypersensitivity to sunlight, and UV light in particular. The deficiency results in skin disorders and cancer predisposition. The disease is caused by a deficiency in nucleotide excision repair.
XP patients cannot excise pyrimidine dimers and other bulky adducts. Mutations occur in one of eight genes called XPA to XPG, all of which encode proteins involved in various stages of nucleotide excision repair. Nucleotide excision repair in eukaryotes proceeds through two major pathways, which are illustrated in FIGURE 14.12.
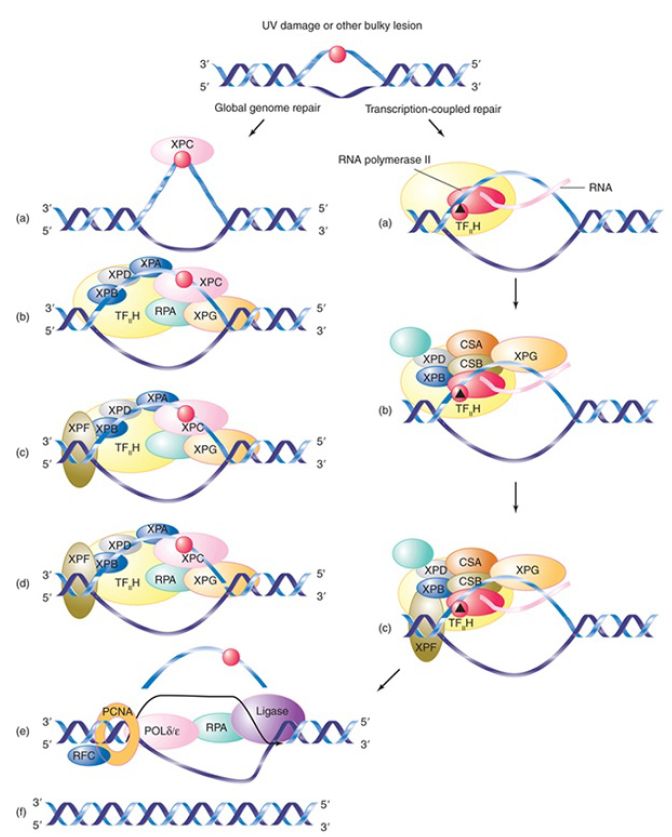
FIGURE 1. Nucleotide excision repair occurs via two major pathways: global genome repair, in which XPC recognizes damage anywhere in the genome, and transcription-coupled repair, in which the transcribed strand of active genes is preferentially repaired and the damage is recognized by an elongating RNA polymerase.
Data from E. C. Friedberg, et al., Nature Rev. Cancer 1 (2001): 22–23.
The major difference between the two pathways is how the damage is initially recognized. In global genome repair (GG-NER), the XPC protein detects the damage and initiates the repair pathway. XPC can recognize damage anywhere in the genome. In mammals, XPC is a component of a lesion-sensing complex that also includes the proteins HR23B and centrin2. XPC also detects distortions that are not repaired by GG-NER (such as small unwound regions of DNA), suggesting other proteins are required to verify the damage bound by XPC. Although XPC recognizes many types of lesions, some types of damage, such as UV-induced cyclobutane pyrimidine dimers (CPDs), are not well recognized by XPC. In this case, the DNA damage-binding (DDB) complex assists in recruiting XPC to this type of damage.
In contrast, transcription-coupled repair (TC-NER), as the name suggests, is responsible for repairing lesions that occur in the transcribed strand of active genes. In this case, the damage is
recognized by RNA polymerase II itself, which stalls when it encounters a bulky lesion. Interestingly, the repair function may require modification or degradation of RNA polymerase. The large subunit of RNA polymerase is degraded when the enzyme stalls at sites of UV damage.
The two pathways eventually merge and use a common set of proteins to effect the repair itself. The strands of DNA are unwound for about 20 bp around the damaged site. This action is performed by the helicase activity of the transcription factor TFII, itself a large complex, which includes the products of two XP genes, XPB and XPD. XPB and XPD are both helicases; the XPB helicase is required for promoter melting during transcription, whereas the XPD helicase performs the unwinding function in NER (though the ATPase activity of XPB is also required during this stage). TF II is already present in a stalled transcription complex; as a result, repair of transcribed strands is extremely efficient compared to repair of nontranscribed regions.
In the next step, cleavages are made on either side of the lesion by endonucleases encoded by the XPG and XPF genes. XPG is related to the endonuclease flap endonuclease 1 (FEN1), which cleaves DNA during the base excision repair pathway . XPF is found as part of a two-protein incision complex with ERCC1, which may assist XPF in binding DNA at the site of incision. Typically, about 25 to 30 nucleotides are excised during NER.
Finally, the single-stranded stretch including the damaged bases can then be replaced by new synthesis, and the final remaining nick is ligated by a complex of ligase 3 and XRCC1.
TF II, particularly the XPB and XPD subunits, plays numerous and complex roles in NER and transcription. The degradation of the large subunit of RNA polymerase II is deficient in cells from patients with Cockayne syndrome, a repair disorder characterized by neurological impairment and growth deficiency, which may also show photosensitivity similar to that of XP, but without the cancer predisposition. Cockayne syndrome can be caused by mutations in either of two genes (CSA and CSB), both of whose products appear to be part of or bound to TF II, and can also be caused by specific mutations in XPB or XPD.
Another disease that can be caused by mutations in XPD is trichothiodystrophy, which has little in common with XP or Cockayne (it is marked by brittle hair and may also include cognitive impairment). All of this marks XPD as a pleiotropic protein, in which different mutations can affect different functions.
In fact, XPD is required for the stability of the TF II complex during transcription, but its helicase activity is not needed during transcription. Mutations that prevent XPD from stabilizing the complex cause trichothiodystrophy. The helicase activity is required for the repair function. Mutations that affect the helicase activity cause the repair deficiency that results in XP or Cockayne syndrome.
In cases where replication encounters a thymine dimer that has not been removed, replication requires DNA polymerase η activity in order to proceed past the dimer. This polymerase is encoded by XPV. This bypass mechanism allows cell division to proceed even in the presence of unrepaired damage, but this is generally a last resort as cells prefer to put a hold on cell division until all damage is repaired.





















