Conjugation Transfers Single-Stranded DNA
KEY CONCEPTS
- Transfer of an F plasmid is initiated when rolling circle replication begins at oriT.
- The formation of a relaxosome initiates transfer into the recipient bacterium.
- The transferred DNA is converted into double-stranded form in the recipient bacterium.
- When an F plasmid is free, conjugation “infects” the recipient bacterium with a copy of the F plasmid.
- When an F plasmid is integrated, conjugation causes transfer of the bacterial chromosome until the process is interrupted by (random) breakage of the contact between donor and recipient bacteria.
Transfer of the F plasmid is initiated at a site called oriT, the origin of transfer, which is located at one end of the transfer region. The transfer process may be initiated when TraM recognizes that a mating pair has formed. TraY then binds near oriT and causes TraI to bind to form the relaxosome in conjunction with host-encoded DNA-binding proteins called integration host factor (IHF). TraI is a relaxase, like ФX174 A protein. TraI nicks oriT at a unique site (called nic), and then forms a covalent link to the 5′ end that has been generated. TraI also catalyzes the unwinding of approximately 200 base pairs (bp) of DNA and remains attached to the DNA 5′ end throughout the conjugation process (this is a helicase activity).
The TraI-bound DNA is then transferred to the T4SS by the coupling protein TraD, where it is exported to the recipient cell. FIGURE 1. shows that the relaxase-bound 5′ end leads the way into the recipient bacterium. The transferred single strand is circularized and a complement strand is synthesized in the recipient bacterium, which as a result is converted to the F-positive state.
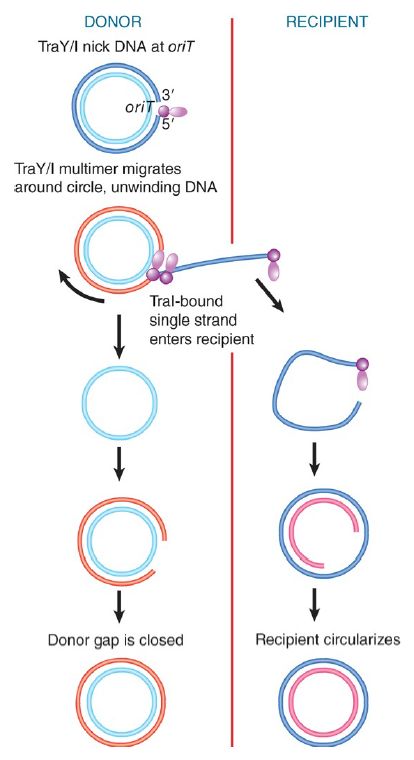
FIGURE 1. Transfer of DNA occurs when the F plasmid is nicked at oriT and a single strand is led by the 5′ end bound to TraI into the recipient. Only one unit length is transferred. Complementary strands are synthesized to the single strand remaining in the donor and to the strand transferred into the recipient.
A complementary strand must be synthesized in the donor bacterium to replace the strand that has been transferred. If this happens concomitantly with the transfer process, the state of the F plasmid will resemble the rolling circle . DNA synthesis could occur instantly, using the freed 3′ end as a starting point. Conjugating DNA usually appears like a rolling circle, but replication as such is not necessary to provide the driving energy, and single-strand transfer is independent of DNA synthesis. Only a single unit length of the F plasmid is transferred to the recipient bacterium. This implies that some feature (perhaps TraI) terminates the process after one revolution, after which the covalent integrity of the F plasmid is restored. TraI might also be involved in recircularization of the transferred DNA to which a complementary strand is then synthesized.
When an integrated F plasmid initiates conjugation, the orientation of transfer is directed away from the transfer region and into the bacterial chromosome. FIGURE 2. shows that, following a short leading sequence of F DNA, bacterial DNA is transferred. The process continues until it is interrupted by the breaking of contacts between the mating bacteria. It takes 100 minutes to transfer the entire bacterial chromosome, and under standard conditions contact is often broken before the completion of transfer.
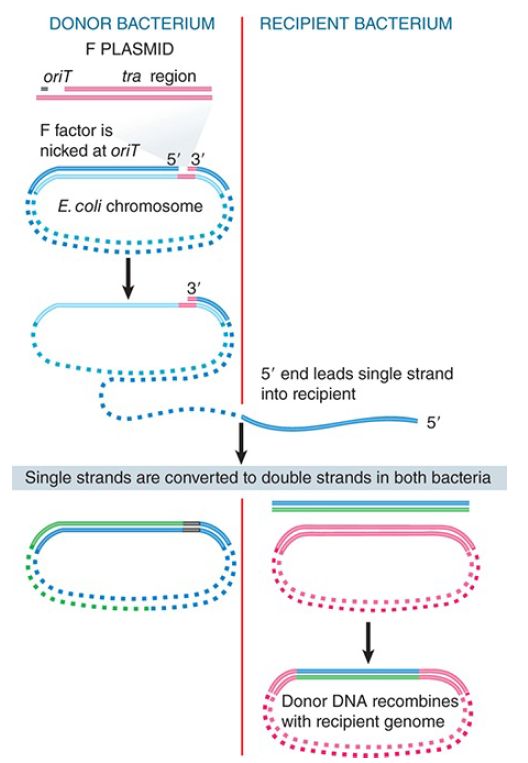
FIGURE 2. Transfer of chromosomal DNA occurs when an integrated F plasmid is nicked at oriT. Transfer of DNA starts with a short sequence of F DNA and continues until prevented by loss of contact between the bacteria.
Donor DNA that enters a recipient bacterium is converted to double-stranded form and may recombine with the recipient chromosome. (Note that two recombination events are required to
insert the donor DNA in order to avoid converting the circular chromosome to a linear form.) Thus, conjugation affords a means to exchange genetic material between bacteria, a contrast to their usual asexual growth (hence the original name Fertility factor or F factor). A strain of E. coli with an integrated F plasmid supports such recombination at relatively high frequencies (compared to strains that lack integrated F plasmids); such strains are described as high-frequency recombination (Hfr). Each position of integration for the F plasmid gives rise to a different Hfr strain, with a characteristic pattern of transferring bacterial markers to a recipient chromosome.
Contact between conjugating bacteria is usually broken before transfer of DNA is complete. As a result, the probability that a region of the bacterial chromosome will be transferred depends on its distance from oriT. Bacterial genes located close to the site of F integration (in the direction of transfer) enter recipient bacteria first, and are therefore found at greater frequencies than those that are located farther away and enter later. This gives rise to a gradient of transfer frequencies around the chromosome, declining from the position of F integration. Marker positions on the donor chromosome can be assayed in terms of the time at which transfer occurs; this gave rise to the standard description of the E. coli chromosome as a map divided into 100 minutes. The map refers to transfer times from a particular Hfr strain; the starting point for the gradient of transfer is different for each Hfr strain because it is determined by the site where the F plasmid has integrated into the bacterial genome.





















