Natural Resistance Genes
The hypersensitive response often results from a specific interaction between a biotrophic pathogen and its host plant. This is known as a ‘gene-for-gene’ interaction between pathogen and host, in which an avirulence gene product from the pathogen is recognised by a resistance gene in the host plant. This recognition leads to induction of hypersensitive defence responses.
Avirulence (Avr) genes encode a variety of polypeptides, some of which may be required for pathogenicity but have then become avirulence factors once they have been detected by the plant. The best characterised Avr genes are Avr4 and Avr9 of the fungal pathogen Cladosporium fulvum. These encode pre-proteins which are processed to mature, extracellular cysteine-rich peptides of 86 and 28 amino acids, respectively. These peptides induce a hypersensitive response in plants which contain the matching resistance genes Cf-4 and Cf-9, respectively.
A series of plant resistance (R) genes active against a range of pathogens have been cloned and characterised. The R genes Cf-4 and Cf-9 encode transmembrane glycoproteins, in which the extracellular portion has characteristic leucine-rich repeats (LRR), a transmembrane domain and a C-terminal cytoplasmic domain. The Avr gene products are recognised by the LRR receptor regions, which results in signal transduction, gene activation and the hypersensitive response.
With the characterisation of R-genes against fungal, bacterial, viral and nematode pathogens, common patterns have emerged. Depending on the site of recognition of the elicitor, the R gene products may either span the plasma membrane and detect the elicitor extracellularly or be located in the cytoplasm for intracellular elicitor recognition. Intracellular recognition would be expected for virus infection, but pathogens growing extracellularly appear to be recognised by the presence of extracellular elicitors or signal molecules from the pathogens that cross the host plasma membrane.
Five classes of R-genes have been proposed (Table 1 and Figure 1), in which common features occur – leucine-rich repeats (LRR), nucleotide binding site (NBS), leucine zippers (LZ), toll and interleukin-
Table 1 Natural resistance genes characterised.
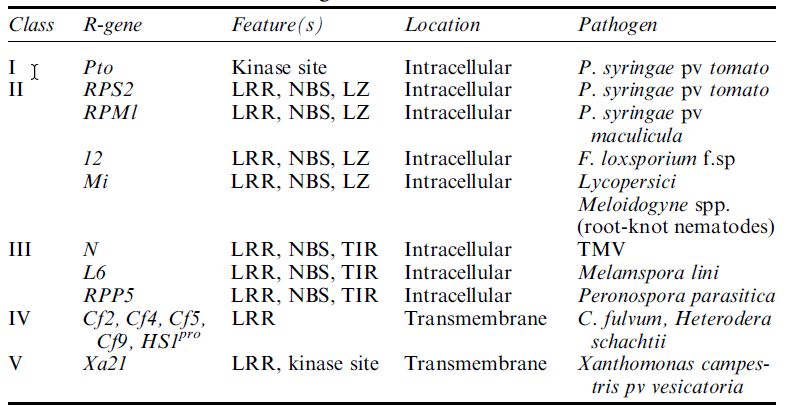
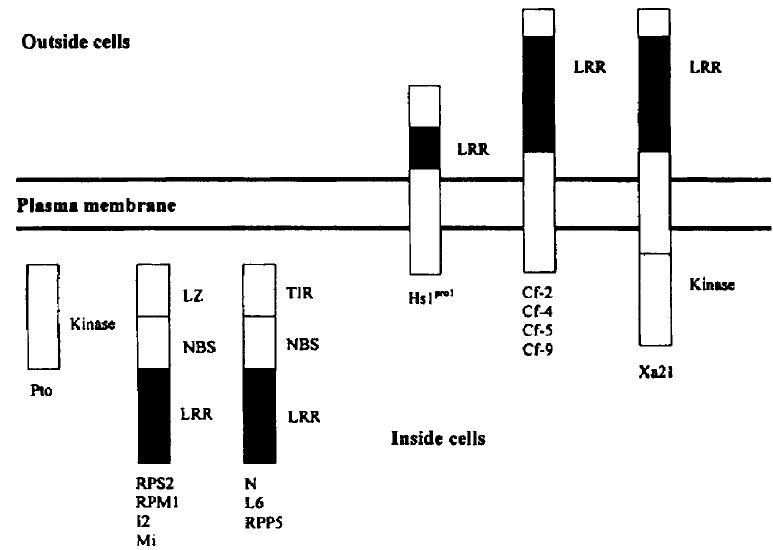
Figure 1 Diagnostic representation of different classes of resistance (R)-genes.
like receptors (TIR) and kinase sites.36 Some of these are shown in Table 10.3. Natural R-genes of this type can be transferred to other plants and, depending on the presence of elicitors/races of pathogen, may confer resistance in other plants. Modification of R-genes to alter the specificity of recognition of elicitors could also be achieved and various schemes have been developed to convert the specific recognition (e.g. of the avr9/ Cf9 system) to a general defence response switched on by damage caused by non-specific pathogens.
As indicated above, most of the R-genes identified so far have common sequence and structural motifs. It is possible to align published sequences and synthesize PCR primers complementary to the conserved sequences and to amplify classes of ‘resistance gene analogues’ (RGAs). RGAs can be mapped and exist in local clusters of related sequences in the genome. This approach can aid identification of natural resistance genes and mapping of RGAs can also provide molecular markers closely linked to known resistance genes.
However, not all R-genes operate via a gene-for-gene mechanism. The HM1 R-gene encodes a reductase which inactivates toxins produced by the fungal pathogen Cochliobulus carbonum and the mlo gene for powdery mildew resistance encodes a negative regulator of cell death.





















