Division Lycophyta
Division Tycophyta contains species that represent a distinct line of evolution out of the first land plants. Lycophytes have lateral sporangia and exarch protosteles, so they may have come from a Zosterophyllum type of ancestor (Fig. 1; Table).


FIGURE 1:A proposed phylogeny of division Lycophyta. The lycopods are believed to have originated from zosterophylls and diversified early; then most became extinct and now only the genus Lycopodium still exists with living species. The selaginellas and Isoetes may have evolved from rhyniophytes that were extremely similar to the zosterophyllophytes and then diversified rapidly into many groups, several of which contained large trees with secondary growth. Most also become extinct, and now only three genera are left.
MORPHOLOGY
The earliest lycophytes were members of the genera Drepanophycus and Baragwanathia (Fig. 2a). They were similar to their presumed ancestors, the zosterophyllophytes, with an important difference: Their enations were large, up to 4 cm long, and they contained a single well-developed trace of vascular tissue. Such enations must have been effective at increasing photosynthesis, and they could be called leaves. However, "leaf1 is an ambiguous term, and enations in the division Lycophyta are called microphylls for clarity (Fig. 2b-e). "Micro-" refers to their evolution from small enations, not to their actual size. In some plants, they were up to 78 cm long. This is not the line of evolution that led to ferns and seed plants; microphylls are not the same as the leaves you are familiar with.
Another important advance over the type of body organization of the Rhynia-Zostero-phyllum type was the evolution of true roots that allowed sporophytes to anchor firmly, absorb efficiently, and thus to grow to tremendous size.

FIGURE 2: (a) Drepanophycus was an early lycophyte that was still small and simple. (b to e) Maophylls in lycophytes are believed to have evolved as enations. Originally they were small, simple flaps of photosynthetic tissue (c). Later they became larger (d) and were vascularized (e).
Many extinct lycophytes such as Lepidodendron, Sigillaria, and Stigmaria had a vascular cambium and secondary growth (Figs. 3 - 4). Their wood looked remarkably like the secondary xylem of pines and other living conifers, having a pith, rays, and elongate tracheids. However, the vascular cambium had one major flaw: Its cells apparently could not undergo radial longitudinal division, so new fusiform initials could not be produced. As the wood grew to a larger circumference, the cambial cells became increasingly wider tangentially (Fig. 4b). No specimen has ever been found with wood more than about 10 cm thick. After that much secondary growth, the cambial cells may have stretched so much that they could no longer function.

FIGURE 3: (a) Reconstruction of Lepidodendron showing a large tree with one distinct trunk (monopodial growth) and a well-developed root system. Leaves (microphylls) were sophisticated, and sporangia occurred in discrete strobili (cones), (b) Sigillaria was a large lycophyte tree whose leaves usually were 1 m or more long. On the branch to the left, leaves have been left undrawn to reveal cons of sporangia.

FIGURE 4: (a) Wood of Sigillaria consisted of large tracheids and superficially resembled the wood of modern conifers. However, it was not as derived in its pitting and other features of its secondary wall, and it was formed by a vascular cambium that could not form new fusiform initials (X 80). (Courtesy of T. Delevoryas, University of Texas) (b) In the vascular cambia of seed plants (top), as the cambium is pushed outward by accumulation of new secondary xylem, its circumference becomes greater, but (uniform initials divide by radial walls, creating new fusiform initials. An old cambium has many more cells than a young cambium. In arborescent lycophytes (bottom), cambial cells could not undergo this division, so an old cambium had no more cells than a young one. The older cells became stretched circumferentially until they were no longer functional; secondary growth then ceased.
HETEROSPORY
Reproduction of lycophytes was also more sophisticated than that of rhyniophytes and zosterophyllophytes in that the sporangia were clustered together in compact groups called cones or strobili (sing.: strobilus), which protected the sporangia (Fig. 5). Although party species remained homosporous, others became heterosporous, having microspores and megaspores that germinated to give rise to distinct microgametophytes and megagametophytes, respectively. Heterospory has special significance in land plants, because it ts a necessary precondition for the evolution of seeds. In the lycophytes Lepidostrobus and Lepidophloios, the megaspore developed into a megagametophyte without enlarging; the megagametophyte existed completely within the wall of the original megaspore, which was up to 10 mm long. Furthermore, the megaspore in some species was retained within the sporophyll, protected by thick-walled cells of the sporangium. This is remarkably similar to ovules and seeds in the seed plants, the most important difference being that in these lycophytes, the sporangium dehisced (much like modern anthers do) and the megaspore wall cracked, exposing the archegonia. Sperm cells could swim to the egg during fertilization.

FIGURE 5: (a) Lycopodium cernuum is a common species that has extensive rhizomes, vertical chlorophyllous shoots, and sporangia clustered into cones. The shoots are actually leaning against and being supported by the surrounding grasses and shrubs. (b) Lycopodium obscurum, with sporangia clustered into cones at the tips of the branches. (Ed Reschke) (c and d) Lycopodium lucidulum is one of several lycopod species in which the sporangia are distributed among the leaves rather than in strobili. (James W. Richardson/VU) (e) Selaginella plants tend to be more delicate than those of Lycopodium and are often overlooked or mistaken for mosses. The leaves usually are very small and delicate and sporangia are inconspicuous. (Dennis Drenner)
The lycophytes are remarkable in that they represent an ancient line of evolution distinct from the seed plants but demonstrate convergent evolution in several characters: leaves, roots, secondary growth, and almost seeds. In the Devonian and Carboniferous Periods, this group dominated the swampy areas of Earth with extensive forests of large trees, but most became extinct; at present the entire division contains only five genera, Lycopodium, Phylloglossum, Selaginella, Isoetes, and Stylites.
EXTANT GENERA
Lycopodium ("ground pine" or "club moss") is fairly common in forests from tropical regions to the arctic (Fig. 5). All living species, about 200, are small herbs with prostrate rhizomes that have true roots and short upright branches. Microphylls are spirally arranged on their stems, and secondary growth never occurs. The sporangia may be arranged in cones or distributed along the shoots (L lucidulum). All Lycopodium species are homosporous, a relictual trait. Spores germinate and grow into bisexual gametophytes that produce both antheridia and archegonia. In some species gametophytes are green and photosynthetic; in others they are subterranean and heterotrophic, nourished by fungi.
Selaginella is less common in temperate North America, and its plants are smaller and easily overlooked (Fig. 23.21e). Probably the best-known species is the resurrection plant, S. lepidophylla, which curls up, turns brown, and appears dead upon drying but uncurls and regreens when moistened. Unlike Lycopodium, Selaginella has the more derived condition of being heterosporous, and the megagametophyte develops inside the megaspore wall . The megaspore is not retained on the sporophyte, however, and is not seedlike. The microgametophytes also develop within the spore wall and consist of a single vegetative cell and an antheridium (Fig. 6b). Many flagellate sperms are produced and then released as the spore wall ruptures. The sperms swim to the archegonia through a film of dew or other available moisture.
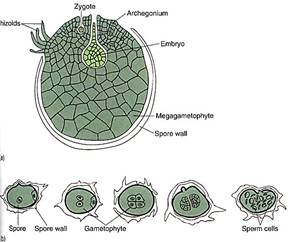

FIGURE 6 :(a) The megagametophyte of some selaginellas develops almost entirely within the megaspore wall. The spore cracks open, exposing the archegonia (megagametangia) and permitting fertilization by swimming sperm cells. (b) The microgametophytes of selaginellas develop within the microspore wall, then liberate motile sperms. (c) Reconstruction of the organelles of a sperm cell of Lycopodium obscurum. These sperms cells are similar to those of mosses (see Fig. 1) and also strongly resemble sperm cells of green algae. Color coding is like that of Figure 1: Blue = nucleus, green = plastid, red = flagella and basal bodies, brown = mitochondrion, yellow = microtubulas that anchor that flagella. (Courtesy of K. Renzaglia, B. Tackett, Jr., and A. Maden, East Tennessee State University)
Selaginellas can be distinguished from lycopodiums by the presence of a small flap of tissue, the ligule, on the upper surface of Selaginella leaves. Although ligules are simple structures and their adaptive advantages are unknown, they are ancient and can be used to distinguish early fossil selaginellas from fossil lycopods.
Isoetes is a genus of about 60 species of small, unusual plants called quillworts which grow in wet, muddy areas that occasionally become dry (Fig. 7). Their body consists of a small corm-like stem that has roots attached below and leaves above. Isoetes is heterosporous like Selaginella, and almost every leaf contains sporangia. Microphylls in this genus also have ligules. Weak cambial growth results in the production of additional cortex parenchyma to the exterior and a type of vascular tissue to the interior; the latter tissue is a mixture of tracheids, sieve elements, and parenchyma. Stylites plants are very similar to those of Isoetes, and several botanists have suggested that the two species of Stylites are really extreme forms of Isoetes (Table).

FIGURE 7:Although Isoetes is a small plant, its short basal corm-like stem has a small amount of secondary growth. (W. H. Hodges/Peter Arnold, Inc.)
The anatomy and morphology preserved in fossil lycophytes indicate many instances of convergent evolution with the seed plants. In both lines, elaborate, efficient leaves, wood, bark, and roots evolved. Sporangia became separated from non-reproductive organs and were grouped together into strobili. In both lycophytes and seed plants, heterospory evolved, as did endosporial development of the megagametophyte. In seed plants, the megaspores were retained in the megasporangium and evolved into seeds; this nearly happened in certain extinct lycophytes. We cannot tell from the fossils if convergent evolution of specialized metabolisms also occurred, but Isoetes does have CAM photosynthesis. It also has a unique means of obtaining its carbon dioxide—absorption by the roots from soil or mud. Its leaves have a thick cuticle but no stomata. We can only wonder what the metabolism of the extinct treelike lycophytes was like. The extinction of so many of the lycophytes is unfortunate; they were complex, sophisticated plants that had many superb adaptations. They probably underwent interesting responses to changes in season, and they must have been able to resist numerous types of pathogenic bacteria, fungi, and insects. Those aspects of their biology will probably remain unknown to us forever.
 الاكثر قراءة في النباتات الوعائية واللاوعائية
الاكثر قراءة في النباتات الوعائية واللاوعائية
 اخر الاخبار
اخر الاخبار
اخبار العتبة العباسية المقدسة




















